
Chapter 10: Vascular Plant Form
One of the striking features of plants is their diversity in form. This is nicely demonstrated with a visit to a greenhouse but can also be seen by looking at outdoor plants, both in native habitats and in gardens. At the same time, all seed plants are fundamentally the same in form and follow a pattern depicted in the Fig. 1 and described as being ‘modular’ and ‘recursive’. What accounts for the apparent diversity in plant form when they are basically put together the same way? There are six key areas of variation that influence the above-ground form of flowering plants and these are listed below and are main sections of this chapter. While the bulk of the chapter deals with flowering plants the form of non-flowering seed plants and vascular plants without seeds is also discussed.
TOPICS
- Flowering plants
- Factors affecting above ground form
- Internode length
- Branching frequency
- Variation in the size, shape and orientation of leaves
- Direction of growth of stems and branches
- Presence or lack of secondary growth
- Loss of parts (leaves and branches)
- Below-ground form
- Factors affecting above ground form
- Form of non-flowering seed plants
- Form of vascular plants without seeds
Factors affecting flowering plant form I: Internode length
-

- Fig. 2a Dandelion rosette. All the leaves are connected to a very short stem.
-

- Fig. 2b Amaryllis has a similar growth pattern although its leaves are more erect
Do dandelions have stems (Fig. 2a)? They must, because they have leaves and leaves are always attached to stems, but dandelion stems certainly are not very evident because they are so very short. You could say that dandelions are ‘vertically challenged’ and as a result, their leaves are very closely clustered, forming what is known as a rosette. Rosettes are a common plant life form and result from the fact that the distance between individual leaves is extremely short, making it appear that there is no stem. While the leaves of dandelion typically run horizontally, parallel to the ground, some plants, like Amaryllis (Fig. 2b) also have very short stems but with leaves that are oriented more vertically. Dandelions live for a number of years and never produce a recognizable stem, although they do produce branches that are flowering stalks. Other plants, including both agricultural crops (e.g., carrots, beets, cabbage, turnip) and common weeds (e.g., mullen, burdock, ‘wild’ carrot, garlic mustard) are biennials. They form a rosette in their first growth season (Fig. 3) and in their second year of growth they ‘bolt’ as the stem produces much longer internodes to produces a more ‘normal-looking’, elongate stem, upon which are born flowers (Fig. 4). A small number of plants, including the one used to make tequila, are a rosette for multiple years, maybe 7-10, before the stem ‘bolts.’


What determines internode length?
Recall that leaves and stem material are produced in the embryonic region called the apical meristem at the very tip of stems. In this region new cells are produced by mitosis and the cells are organized as new stem material and as leaf primordia, outgrowths from the stem that grow and produce the structures that we know of as leaves. In dandelions and other rosette plants, the distance between leaves is very short. As mentioned above, internode length may vary with age, or more specifically vary with environmental factors that coordinate with age, in which case plant form may drastically change with age (see images of mullen, above). Internodes produced in the first season of growth are very short, those in the second season of growth, after the plant has been exposed to a cold period, are a great deal longer.

What are the consequences of producing rosettes as opposed to stems with leaves more widely spaced (Fig. 4, 6)? One factor is microclimate, rosette leaves, if they are close to the ground, may be in a very different environment that leaves that are elevated. A second factor is self-shading of leaves, although this is influenced by several other factors including the size of the leaves and their distribution on the stem. Also, plants with short stems and clustered leaves are also more likely to be shaded from above because the stem is not carrying leaves upwards and potentially above the neighboring plants. At the same time, growing upward exposes the plant to a number of problems that are not present if it stays low to the ground. One is mechanical. Growing upward requires stems to be able to resist the combined effects of gravity and wind. Another problem is that an elevated shoot apical meristem becomes more apparent and vulnerable, at least to some herbivores. The loss of the terminal apical meristem is particularly damaging because it eliminates the source of growth, although growth may continue by the activation/stimulation of lateral buds. The accessibility of the stem tip depends on the herbivore—something big, and especially something big with a big nose (think deer!) might have a hard time nipping the tip of a dandelion, but have no difficulty nipping the tip of a tomato plant. But for smaller herbivores like insects, stem tips are available for both short and tall plants.

 Fig. 6 Impact of internode distance. The top image of a Diffenbachia stem shows the widely spaced-out leaf scars about 3 cm apart. The bottom image of cabbage shows a very distinctive rosette form.
Fig. 6 Impact of internode distance. The top image of a Diffenbachia stem shows the widely spaced-out leaf scars about 3 cm apart. The bottom image of cabbage shows a very distinctive rosette form.Factors affecting flowering plant form II: Branching frequency
Note the images of Diffenbachia and cabbage (Fig 6). Like dandelion, they have no branches. ‘Typical’ plants with modular growth and a dendritic form have branches and the branches have branches. This repetitive pattern is one of the familiar aspects of plant form that makes them so appealing as an artistic model. However, some plants, like dandelion, Diffenbachia and cabbage, have no branches, there is just one axis to the plant, and this gives them, and other plants, a distinct look.
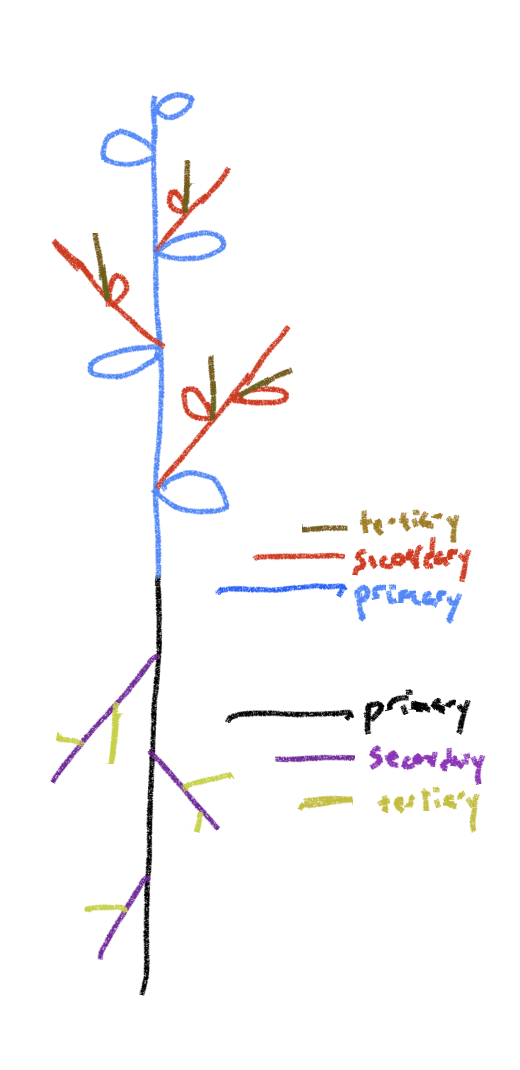
Recall that branches result from the fact that as the shoot apical meristem elongates and produces embryonic leaves it also produces a potential branch, called a branch primordium or bud primordium in the ‘axil’ of each leaf, placed just above where the leaf attaches to the stem. A branch primordium is a replica of the shoot apex, an embryonic shoot capable of elongating and producing a shoot with leaves upon it. All that needs to happen to produce a branch is that the branch primordium be stimulated to start growing. Branch upon branch upon branch can be produced as each shoot in turn produces leaves that have branch shoots appear from their base. This does not happen in dandelion and in many other plants, both rosette and non-rosette. In such plants, growth occurs solely from the original shoot, although many of them will form branches if the original shoot apical meristem is removed. In dandelion, although branch primordia are produced, they are only activated to grow after being transformed into flower primodia (technically inflorescence primordia). Thus, when activated, they produce not a branch but a leafless dandelion ‘flower’ (technically it is an inflorescence, a cluster of flowers, not a single flower). This type of growth pattern is also seen in Aloe and a number of other rosette plants.
Frequently one may see a cluster of dandelion plants together, which might appear like a group of branches. However, these are usually formed when the root of a dandelion is broken off (usually as the result of someone trying to pull it up). The root that remains is stimulated to form new (adventitious) shoots. Often what happens is that multiple shoots are produced by a single root, forming in a cluster of stems. Each of them originated separately from the root tissue, so what you see is not a cluster of branches but instead a cluster of adventitious shoots.
Often branches don’t look like branches
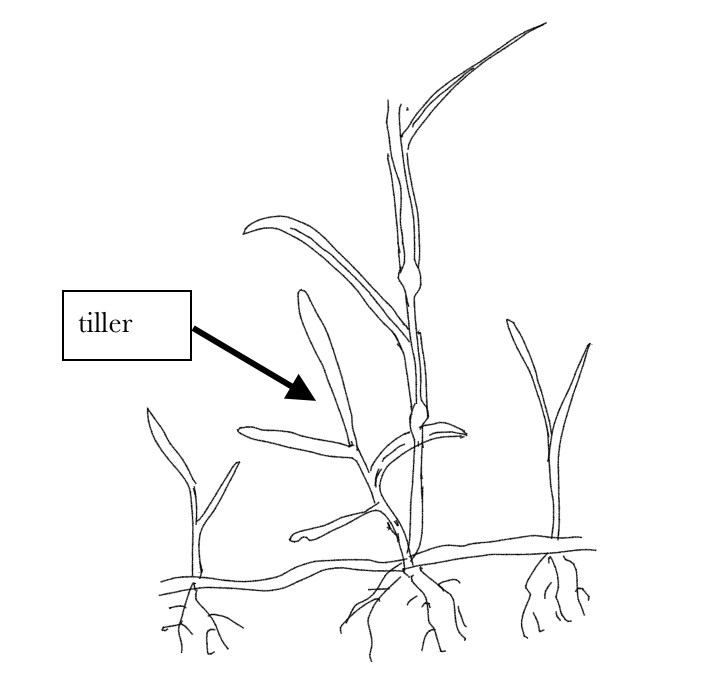
In grasses and many other plants, the branches often aren’t recognized as such. Branches often appear at the base of the plant when branch (bud) primordia of the lowest leaves are stimulated to grow in one of three ways (Fig. 7 and 8): (1) extending vertically, producing a leafy stem called a tiller in grasses (on non-grasses this sometimes called a ‘sucker’ or a ‘pup’) that is basically the same in structure and function as the original stem; (2) extending horizontally and above-ground, producing only rudimentary leaves with very reduced blades and elongate sheaths. These horizontal stems (branches) are called stolons; (3) extending horizontally but below ground, again producing only rudimentary leaves. These horizontal underground stems (branches) are called rhizomes. Stolons and rhizomes are sometimes considered a means of reproduction; they can also be thought of as providing mobility. Eventually both produce vertical, photosynthetic stems that originate from the bud primordia associated with the rudimentary leaves. A number of plants (e.g., goldenrods, asters) that appear to exist as vertical stems are actually a group of vertical branches all connected to a below-ground stem (rhizome).

Factors affecting flowering plant form III: Variation in the size, shape and orientation of leaves

In contrast to dandelions, which look like they don’t have a stem when they actually do, grasses (and a number of other plants, including bananas) possess something that looks like a stem but actually isn’t. The ‘trunk’ of a banana plant is actually a cluster of leaf bases tightly wound (Fig. 9) around each other and called a ‘pseudostem’ because it has the appearance of a stem. Bananas and grasses are monocots, a group that includes orchids and a number of other flowering plants. One feature commonly found in monocots, and specifically in banana and grasses, is a leaf with two main parts (Fig. 10). One part is called the blade; this is typically flattened and extends outward from the plant and is often roughly horizontal to the ground. This is the part that most people recognize as a leaf. If you follow the blade back to the rest of the plant, it abruptly narrows (a great deal in banana, less so in grasses) to a section of the leaf that is vertically oriented and cylindric. This part of the leaf is called the sheath. Early in the growth of both grasses and bananas the structure that looks like a stem is actually just multiple leaf sheaths extending upward from a very short stem, like a dandelion stem.
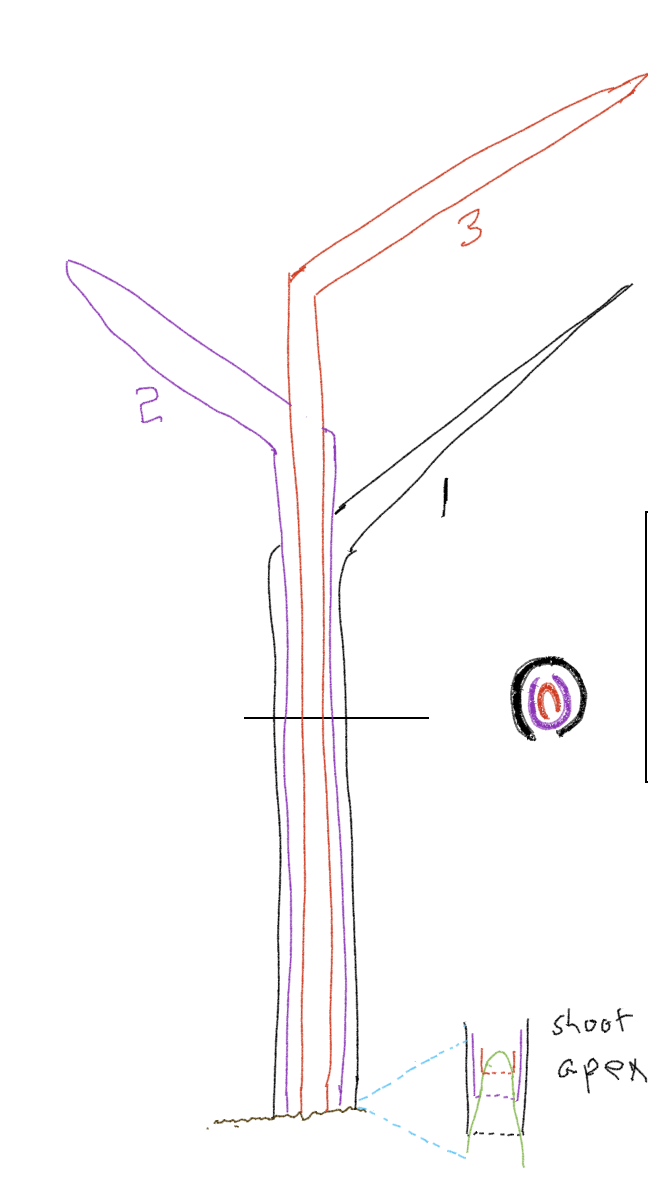
While dandelion leaves are generally horizontally displayed, in grasses the bases of the leaves (i.e., the sheaths) are vertical. The more recently produced leaf sheaths are located inside of older ones and they collectively form a structure that seems like a stem. In grasses and bananas, the stem eventually elongates, the result of very large increase internode length of the youngest leaves. If one were to measure the internode lengths of the first 10 leaves of a grass they might be (in mm and starting with the first leaf produced): 1, 1, 1, 1, 1, 10, 50, 100, 150, 200. After producing six leaves the stem is 6 mm in height, after producing four more leaves it is 506 mm in height. The stem ‘telescopes’ upward inside of the sheaths, and often exposes the uppermost nodes, the actual point attachment of leaves to the stem. When the stem of a grass or a banana elongates, it signals the end of the life of that stem and its associated leaves. An inflorescence is produced at the end of the stem, with flowers that develop into fruits. Nutrients are mobilized from the leaves to the developing seeds and fruits. As noted above, many grasses and bananas branch from the base and this may perpetuate the organism even if the original stem flowers, senesces and dies.
Dandelion, grass and banana leaves just scratch the surface of variation in leaf form. Palm leaves may be up to 25 m in length and duckweed leaves are about 3 mm (Fig. 11, 12). Most leaves are flat but some are thick (succulent) and may be round and bead like (Fig. 13). Some leaves are appressed to the stem (Fig. 14). Leaves can be lobed and toothed in a variety of ways. Many leaves are dissected into parts sometimes in such ways that it isn’t obvious whether one is looking at a leaf or a leaflet (Fig. 15).





Factors affecting flowering plant form IV: The direction of growth of stems and branches: vertical, horizontal or something in between
Although most familiar plants have a main stem and branches that grow upwards, this is not always the case. As mentioned earlier, branches may be hidden below ground because they do not grow vertically but instead extend horizontally below the ground. As a result, multiple stems rising from the ground may not be separate plants but actually be branches that extend vertically from a main stem that is running horizontally below ground. While most familiar plants have stems that grow upwards, the exact angles vary and some run horizontally, producing what is known a ‘prostrate’ growth form seen in partridge berry and strawberry below (Fig. 16-17)


Some plants (pin oak) have branches that extend roughly 90 degrees from a vertically oriented stem, i.e., they extend horizontally. Other plants have branches that run more vertically, perhaps at an angle of 45 degrees from the main stem.
It is also quite common that orientation changes, generally in a controlled manner. In the example of grasses or banana described above the original stem is vertical but 1st order branches may run horizontally (if they are rhizomes or stolons) and third order branches (i.e., branches off of the stolon/rhizome) again run vertically. Thus plants vary in the way that the original stem and its branches are oriented, both within a plant and between different species.
Factors affecting flowering plant form V: whether or not the stems and branches exhibit secondary (generally woody) growth
Secondary growth allows plants to become wider and taller. While most people associate woodiness with trees, two other plant forms are woody: ‘Shrubs’ have no rigid definition but are tree-like but show limited growth in height but abundant branching (Fig. 18). Most vines (lianas) are woody and have an interesting cellular nature that lacks sclerenchyma fibers and consists almost entirely of water conducting cells with relatively thin secondary walls. This is possible because they utilize the structure of other plants to provide support. Consequently vine wood has a low density and rigidity when compared to tree wood.

Factors affecting flowering plant form VI: shedding of plant parts
A final factor that impacts the ‘look’ of plants is the shedding of plant parts. Most plants discard pieces as they grow. Generally, the discarded parts are leaves but branches may also be shed (Fig. 19-20). The loss of parts may be synchronous, as it is with most deciduous trees, but other plants may shed parts continuously. Plant structure is dynamic and parts are not solely being added, they are generally also being removed. These changes are often not noticed, except by gardeners/landscapers, because the changes are relatively slow, the new growth looks just like the old growth and the pieces shed are matched by pieces added. For most plants both the growth and the shedding are usually sporadic, occurring in spurts with inactivity between them.


Below-ground form
Root systems also have a variety of forms but, obviously, it is harder to see the form of root systems. One distinction that is often cited is the distinction between a taproot system, where the embryonic root (the one formed in the seed) persists and becomes the main trunk of the root system that connects with the stem. Note that a carrot is perhaps not the best example of this, even though it is often used, because in carrot the root is performing a carbohydrate storage function, something that does not have to be the case for plants with taproots. In contrast to taproots, many plants have a root system that forms from adventitious roots that are produced off of stem/rhizome/stolon and do not have a dominant, primary root. Fibrous root systems may only have a few levels of branching whereas taproots typically have more, especially as they age. The primary function of roots is to acquire water and the pattern of water distribution is significant to root system form, specifically lateral spread vs. vertical spread. Alfalfa roots are known to penetrate 15 meters down in order to obtain water. On the other hand, most cactus roots spread extensively but do not penetrate deep in the soil. Cactus also commonly produce roots quickly after a rain and then rapidly loose them (i.e., they senesce and die) as the soil dries. All plants, not just cactus, shed root structure; similar to the situation aboveground, root structure is dynamic, with pieces continually being added and shed.
Form of seed plants without flowers
There are four groups of seed plants that lack flowers. Representatives of most of these groups are present in the ‘Organisms to Know’ section and are highly in blue.
The ginkgophyte group only has a single species, ginkgo, which in form is typical of a deciduous (angiosperm) tree (Fig. 21), except that the leaves are unusual, being fan shaped with bifurcating parallel veins. Ginkgo also has ‘spur shoots’ (‘short shoots’), branches with a very short internode distance and consequently leaves clustered very close together. Spur shoots make identification of ginkgo easy during the winter months.

A second group are the conifers, including pines, hemlock, redwoods and juniper. For most, but not all, conifers the leaves are present on the plant for more than a year, making the tree ‘evergreen’. Many conifers have needle-like leaves although some have small, scale-like leaves that overlap each other, producing what are essentially photosynthetic twigs which are shed as a unit after senescence. Most conifers have tree-like forms (Fig. 22) with a fairly rigid branching pattern that produces the characteristic ‘Christmas tree’ shape. There are a few conifers that are shrubs, primarily in the yew-family and some in the cedar family.

A third group of non-flowering seed plants are the cycads, most of whom have a distinct form, generally with an unbranched, short stem axis and relatively long pinnately compound leaves (Fig. 23). These stem and leaf features are found in two other groups with which cycads may be confused: tree ferns (see below) and palms (a group of flowering plants). Distinguishing between the three groups is easy if reproductive structures are present: palms produce flowers and seeds; cycads have no flowers but produce seeds in a type of cone; and tree ferns produce no seeds but have clusters of spores visible on the leaves).

The fourth group of seed plants lacking flowers, the Gnetum group, has only three genera, each with a very distinctive forms Welwitchia (there is only one species and it lives in southern Africa) is by far the most bizarre of the group (Fig. 24). It produces only two leaves which grow from the base and elongate to form long, strap-like leaves that often shred longitudinally so it appears that there are more than two leaves. The apical meristem dies after these two leaves are produced and hence there is no stem. Reproductive structures are produced by the ‘crown’ at the base of the leaves and plants are either male or female.

A second genus in the Gnetophyta groups is Ephedra (roughly 70 species), whose members are highly branched shrubs with photosynthetic stems and small leaves that are soon shed (Fig. 25).

The final Gnetophyte genus is Gnetum (roughly 40 species) which has species that look the most ‘normal’ (i.e., like many flowering plants) of the Gnetophyte group, with ‘normal’ leaves (determinate structures with a petiole and a flattened blade) occurring on forms that could be described as small trees, shrubs or vines.

Form of vascular plants without seeds
We will consider three groups of plants that possess vascular tissue but do not produce seeds. Again, note that representatives of these groups are described in the ‘organisms to know’ section.
The least diverse and easiest to identify are the horsetails (Equisetum). There are less than 20 species and all look similar and have a distinctive form (Fig. 26-27). They have below-ground rhizomes that produce erect ribbed and ‘jointed’ stems with whorls of minute, scale-like leaves occurring at the joints. Some species vary from forms with and without whorls of branches, while other species are strictly unbranched or strictly branched. Aall living species show no secondary growth bit relatives in the Paleozoic era, 300 million years ago, had tree-like forms with secondary growth and stems up to 30 m high.

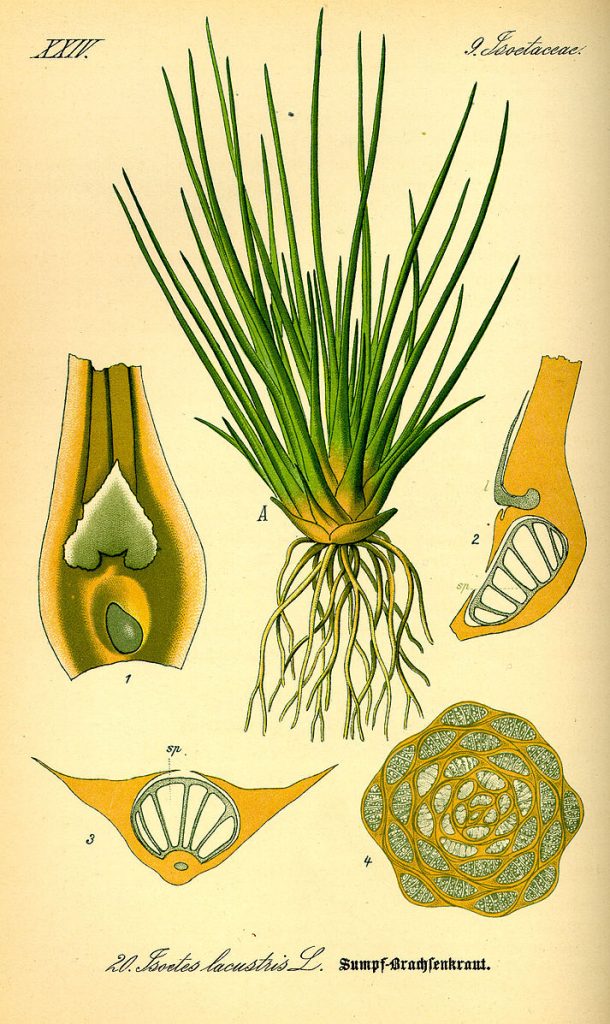
Most members of the clubmoss group have a typical plant ‘stem with leaves’ structure. The stems and roots are vascularized and the leaves have a single vascular trace (vein). Probably because of this, leaves are small and often appressed to the stem, giving the plants a ‘moss’ look, although the plants and leaves are bigger than almost all mosses. Although representatives of the group used to possess secondary growth and tree-like forms, all forms present now are small, usually less than 20 cm tall, and non-woody (Fig. 28-29). Most species have stolons or rhizomes from which vertical branches appear. These may or may not branch. One genus in the group (Isoetes, the quillworts) has a very different form. It is an aquatic, unbranched plant with very short internodes, forming rosettes with awl-shaped leaves that may be 10 cm or more in length (Fig. 30).


The fern group is by far the largest group of vascular plants without seeds and it is the group with the most diversity in form, although the majority of ferns seen in north temperate habits are generally uniform in structure with an underground, woody (i.e. showing secondary growth) rhizome from which leaves arise (see sensitive fern, wood fern). Thus, when you see a fern you are generally seeing a group of leaves (fronds) that originate in a below-ground woody rhizome. Some species have rhizomes that are short and vertical and produce a circular cluster of leaves resulting in an urn-shaped display (Fig. 31). Other species have rhizomes that run strictly horizontally and producing a more uniform ‘patch’ of ferns (Fig. 32). the leaves can be more or less dense depending upon the pattern of leaf production along the rhizome.


In the tropics there are some ferns that are tree-like with an above ground stem vertically oriented stem that may be 2 m in height producing plants that superficially resemble palms and cycads. There also are aquatic ferns that are small and float on the surface of water (Fig. 33), comparable to growth of duckweed, which is a flowering plant. A final example of the diversity in fern form is Marsilea, a fern that resembles four-leaf clovers (Fig. 34) and has a very interesting pattern of sex discussed in Chapter 13.


Further Reading and Viewing
- Fern Structure by the U.S. Forest Service. An excellent description of fern form.
- “Intriguing Lime-Green Blobs Appear In The Andes Mountains. Are They Alive?” by Robert Krulwich. Llareta, an amazing plant.
- “Maximum rooting depth of vegetation types at the global scale” by J. Canadell et al.
Media Attributions
- Mullen Weed © Wilhelm Zimmerling PAR is licensed under a CC BY-SA (Attribution ShareAlike) license
- Mullen Weed © Sanja565658 is licensed under a CC BY-SA (Attribution ShareAlike) license
- Turnips © thebittenword.com is licensed under a CC BY (Attribution) license
- Witte_kool © Rasbak is licensed under a CC BY-SA (Attribution ShareAlike) license
- Rhizome
- Duckweed © Stefan.lefnaer is licensed under a CC BY-SA (Attribution ShareAlike) license
- Cocos nucifera © Ahoerstemeier is licensed under a CC BY-SA (Attribution ShareAlike) license
- Seneciorowleyanus © Stickpen is licensed under a Public Domain license
- Tamarisk_(Tamarix_gallica) © Phil Sellens is licensed under a CC BY (Attribution) license
- Box elder leaf
- leaves
- Fragaria vesca (habitus) © Hans Hillewaert is licensed under a CC BY-SA (Attribution ShareAlike) license
- Artemisia tridentata wyomingensis © Matt Lavin is licensed under a CC BY-SA (Attribution ShareAlike) license
- Trees in the woods
- Tree with several broad trunks
- Landau Ginkgo Goethepark © Rudolf Wild is licensed under a CC BY-SA (Attribution ShareAlike) license
- Picea pungens glauca © Crusier is licensed under a CC BY (Attribution) license
- Voi cycad (Encephalartos kisambo) © Mokkie is licensed under a CC BY-SA (Attribution ShareAlike) license
- Welwitschia mirabilis © Joachim Beyenbach is licensed under a CC BY-SA (Attribution ShareAlike) license
- Ephedra viridis © Stan Shebs is licensed under a CC BY-SA (Attribution ShareAlike) license
- Equisetum hyemale subsp affine © Matt Lavin is licensed under a CC BY-SA (Attribution ShareAlike) license
- Equisetum palustre illustration © Carl Axel Magnus Lindman is licensed under a Public Domain license
- common forest clubmoss © Kirisame is licensed under a CC BY-SA (Attribution ShareAlike) license
- Lycopodium complanatum © Franz Xaver is licensed under a CC BY-SA (Attribution ShareAlike) license
- Illustration Isoetes lacustris © Prof. Dr. Otto Wilhelm Thomé is licensed under a Public Domain license
- Cinnamon Fern cluster © homeredwardprice is licensed under a CC BY (Attribution) license
- Fern Carpet © Nicholas A. Tonelli is licensed under a CC BY (Attribution) license
- Azolla Bed © Komal Sambhudas is licensed under a CC BY-SA (Attribution ShareAlike) license
- Two fern illustration © Dr. Otto Wilhelm Thomé is licensed under a Public Domain license

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_(3963842416).jpg){kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}