
Chapter 28: Agriculture

What is agriculture? It is an activity whereby an organism actively cultures/cultivates/cares for other organisms with the intent of somehow utilizing these organisms directly or indirectly. While we are focused on the agricultural interactions involving humans, other organisms, specifically numerous ant species, carry out agriculture, culturing fungi, aphids and other organisms. Agriculture generally involves the ‘domestication’, a genetic modification of organisms, both plants and animals, allowing them to be better utilized.
Agriculture involving humans and plants is certainly one of the most significant biotic interactions, an activity that influences vast expanses of land, roughly 12% of the earth’s land surface. Although there is much less agricultural activity in aquatic/marine systems, there is some, and secondary effects from terrestrial activities, e.g. fertilizer run-off, also has an impact. Like all interactions, agriculture involves modifications of both partners. And while we will focus on the non-human component, generally flowering plants, it is important to keep in mind that agriculture has brought about tremendous changes in humans, with probably the most significant ones coming about as agriculture originated and humans altered their patterns of activity, movement and social interactions. It is significant to note that relative to the existence of humans, agriculture is a recent innovation, starting roughly 10,000 years ago. For most of their 200,000 year existence, humans were not involved with agriculture, although they certainly did influence specific plants by their activities. It is also significant that agriculture apparently developed separately in multiple regions. Authorities now list eleven sites of agricultural origin, including the Middle East, Africa, the Far East, and North, South and Central America. In each of these regions distinct species were utilized. The table below lists some of the earliest crops along with their region of origin:
Table 1.
|
crops |
site of domestication |
|
wheat, peas, lentils, flax, figs, chickpeas |
Middle East |
|
potato, tomato, pepper |
South America |
|
corn, squash, beans |
Mesoamerica |
|
banana, sugarcane, taro |
New Guinea |
|
rice, mung beans, soy beans |
China |
|
millet, sorghum, |
Africa |
|
eggplant, mungo bean, pigeon pea |
India |
The majority of crops, and certainly the crops associated with the origin of agriculture, were wild plants that humans had already discovered to have desirable features (most commonly features related to eating but sometimes the features were mechanical, e.g. cotton, or chemical, e.g. a waterproof latex from tree bark). The next step was the appreciation that the plant could yield more product if it were somehow cared for, e.g. planted, pruned, etc. For most crops, the practice of collecting and planting seeds was crucial to crop development. Through time, many plant species were genetically modified by humans through selection and planting of seeds from plants with particularly desirable traits, e.g. large seeds. Genetic modification of agricultural organisms (Darwin called this ‘artificial selection’) was most rapid when dealing with annual plants that could be selected for yearly. This chapter examines the histories of several crops that reveal interesting biological features.
TOPICS
- How do crops come to be?
- Wheat
- Strawberry
- Naval oranges
- Banana
- Corn
- Brassica
- Generating variability
Wheat

The plant known as ‘ wheat’ actually includes three distinct species in the Triticum genus: einkhorn wheat, emmer wheat, and bread wheat. Each of these species has wild relatives that may be considered separate species or may be lumped with the domesticated form. These species are related evolutionarily in what is known as a polyploid series. Einkhorn has a diploid chromosome number of 14 and produces haploid gametes with seven chromosomes. Emmer wheat has 28 chromosomes, 14 of which came from Einkhorn wheat and 14 of which came from a related, but separate genus (Aegilops, although some workers have lumped Aegilops and Triticum). Bread wheat has 42 chromosomes, 28 of which come from emmer wheat and 14 from another species of Aegilops.
Polyploidy is common in plants and is an aspect in the history of multiple crops. It can come about several ways, the most common being a hybridization event where gametes of two separate species are combined. Viable gametes cannot be produced in the hybrid because meiosis is thwarted by the fact that there are no homologous chromosomes to pair, the chromosomes from one parent do not have ‘matches’ because the second parent was a different species with different chromosomes. Consequently, the hybrid is sterile; this is what causes the two parents to be considered separate species, they are reproductively isolated.
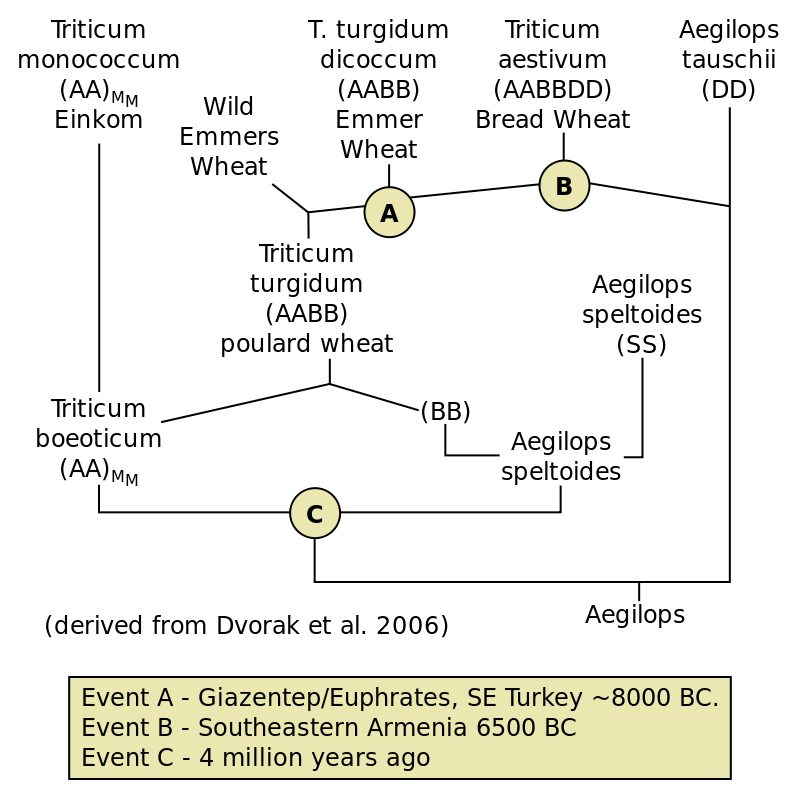
However, hybrid sterility can be overcome in several different ways. One is to produce functional gametes (i.e. cells that can fuse with other cells) without going through meiosis, i.e. to produce ‘unreduced gametes’. If these ‘unreduced gametes’ find each other and fuse then a new species is created, one that has twice the chromosome number of its either of its parents. In the case of emmer wheat the new species has a diploid chromosome number of 28, 14 (seven pairs) coming from einkhorn wheat and 14 (seven pairs) coming from Aegilops. Another possibility is that the cells within the hybrid replicate their chromosomes (mitosis) but the cell does not divide, leaving a cell with double the number of chromosomes of its parents, and significantly, producing a cell that has a match for each chromosome. Such a cell (or derivatives of this cell) could go through meiosis because it does have pairs of chromosomes. In both of these situations a new ‘polyploid’ species like emmer wheat is produced. Polyploidy is also discussed in Chapter 31.
Bread wheat was produced by the same polyploid mechanism following hybridization, this time between emmer wheat and another species of Aegilops. Hence, bread wheat possesses three genomes (sets of chromosomes), one from einkhorn wheat and two from two different species of Aegilops. Each set consists of seven chromosomes, thus dipoid cells of bread wheat have 42 chromosomes, two copies of each of the three sets of chromosomes. The origin of bread wheat is quite recent, less than 10,000 years ago, and after both einkhorn and emmer wheat had been domesticated.
Strawberry

The strawberries found in grocery stores have a very interesting heritage involving Chile, eastern North America and France. Strawberry is in the genus Fragaria and there are over 20 species occurring primarily in temperate regions of North America, Europe and the Far East. Most of these species have seen very limited agricultural utilization but are harvested in the wild. Although it has very limited commercial production, one European species, F. vesca, especially some clones with particularly large fruits, has been cultivated for over 500 years, primarily in parts of Turkey. A South American species with white fruits, F. chiloensis, native to the west coast of North and South America was cultivated by native tribes in what is now Chile. In the early 1800’s six specimens of F. chiloensis were transported to France and propagated in several gardens alongside specimens of F. virginiana, a North American species. The two species hybridized, forming what is called ‘garden strawberry’, Fragaria x ananassa (the ‘x’ in the name indicates that it is a hybrid). It is this hybrid that is now widely cultivated throughout the world, generally being propagated through cuttings.
Navel oranges
Another crop with a ‘chance’ origin is the navel orange. It occurred as a ‘sport’, a mutant branch, on an orange tree growing in Brazil. Remember that branches originate from lateral buds, meristematic tissue left behind by the elongating shoot apical meristem. Occasionally, some of the lateral bud meristems possess mutations that cause them to produce a branch that grows abnormally or one that produces leaves that are unusual (a common manifestation is a branch that produces variegated leaves, leaves that are not uniformly green but are colored a variety of ways). In the case of the navel orange the branch was unusual because its flowers, which normally occur singly, occured as pairs, with a second flower produced very close to the ‘normal’ flower. The proximity of the two flowers is manifested in an altered fruit development, producing not two distinct fruits but rather a single fruit with another fruit inside it. This is what produces the navel for which the plant is named. If one peels and opens up a navel orange, the second fruit is very evident at navel end. This fruit does not possess a skin and is much smaller than the normal fruit, typically less than an inch in diameter. But, like the normal fruit, it is composed of wedge-shaped sections. A second abnormality of navel oranges is that the pollen is sterile and consequently cannot fertilize flowers. Because of this, the fruits produced by navel oranges are seedless.

Navel oranges are perpetuated by cuttings, and all the navel oranges grown world-side are ultimately derived from to the original branch produced on the tree in Brazil. A cutting can be rooted to form a naval orange tree but are more commonly cuttings are grafted on to an existing root stock. A variety of species, generally close relatives, can be used as rootstocks and rootstocks can be selected for, and developed for, favorable characteristics, e.g. temperature, drought and pathogen tolerance.
Banana
Unlike naval oranges which are propagated by grafting branches (called scions) on to a variety of rootstocks, bananas are propagated by the cloning of whole plants, i.e. taking a cutting, usually a branch, and having it form roots. Cloning from cuttings is much less ‘technological’ than grafting and the utilization of cuttings as an agricultural technique goes back to the time when agriculture was developing. There are a large number of banana varieties and it is thought most appeared in the wild and were selected because of favorable features, in particular large fruits that lack seeds. Obviously a lack of seeds makes propagation by seed impossible, but cloning allows these favorable characteristics be perpetuated. Modern studies indicate that the many banana varieties are derived

from two species and polyploid derivatives of these species. Most bananas grown commercially are triploid, with a genetic constitution of AAB meaning that they possess two chromosome sets of one type and one chromosome set from another. Such a triploid may be the result of a tetraploid (AAAA, that produces gametes that are AA) hybridizing with a (normal) diploid (BB, that produces gametes that are B). The offspring of this cross are sterile because meiosis is impossible, but cloning allows the plant to be perpetuated. Moreover, the sterility brought about by hybridization has created a favorable feature — no seeds. Triploids may also arise by the the union of an unreduced gamete from a diploid species (the species is AA but produces gametes that are AA instead of A) combining with a normal gamete from another species, e.g. a diploid species BB producing haploid gametes, B. The offspring is triploid, AAB, and sterile.
For most plant species the production of fruits is a consequence of the production of seeds, with the initiation of seed development triggering the initiation of fruit development. The production of fruits when seeds are not developing at all, or when seeds are initiated but soon aborted, is called parthenocarpy. It is generally considered to be evolutionarily unfavorable since the basic function of fruits is to promote seed dispersal and producing fruits without seeds is a waste of resources. Parthenocarpy can appear ‘spontaneously’, but it generally will not be perpetuated, unless of course it is selected by (early) agriculturalists. More recently, parthenocarpy may be developed in breeding programs. An example is the seedless watermelon, which, like most bananas, is a sterile triploid, but it was produced by plant breeders crossing a tetraploid plant with a (normal) diploid plant. Seedless watermelons do have seeds but they are small and not fully developed. Seedless watermelons do require pollination to initiate the seeds but they soon abort. In some crops (e.g. some varieties of tomato) parthenocarpic varieties produce fruits even if pollination is lacking.
Corn
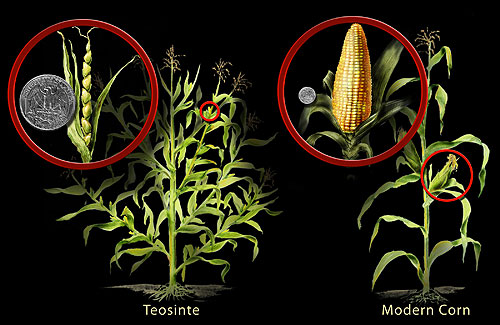
Corn’s origin as a crop has been a mystery because there had been no obvious ancestor, i.e. a species that looks like corn and could be considered to be something that primitive agriculturalists might manipulate to produce the entity that we know as corn. Research over the last 75 years has revealed that there is a close relative, called teosinte, but its proximity to corn is obscured by the fact that it ‘looks’, i.e. has a morphology, that is substantially different from corn. Studies have revealed that corn and teosinte are actually the same species — they readily interbreed. The morphological difference between the two are actually the result of changes in only a few (less than 10) genes. It is now thought that selection on teosinte developed the variety (subspecies) that we now consider to be corn, a plant that unlike teosinte: does not branch, has much larger ‘ears’ (clusters of female flowers), and has seeds that are not enclosed in a rigid container but are relatively easily removed, allowing easier access to an edible structure.
Corn was responsible for dramatic changes in agriculture, in particular the development of seed companies, commercial entities that provide seed to individual farmers. Up until early in the 20th century most farmers provided seeds for themselves by storing seed from the previous crop. When storing seed for the next year’s crop, most farmers, passively or actively, selected for increased yield by saving seed from plants that were disease resistant, pest resistant and generally higher yielding. Early in the 20th century agriculture changed drastically in a number of ways, one of them being that farmers started to purchase seed from companies because they could provide seed that was better than what farmers had on hand. This better seed was the result of agricultural research occurring both in ‘land grant colleges’ (who were charged with improving agriculture) and also in private companies when it was realized that that money could be made by supplying seeds to farmers. Corn was instrumental to this process because it is amenable to producing what is known as ‘hybrid seed’, discussed further in Chapter 31.
The amazing Brassica group

Plants in the genus Brassica reflect a wide variety of interesting topics that intersect agriculture and biology.
Brassica is a genus in the mustard family that has a long history of cultivation and manipulation that has allowed the group to be widely utilized in numerous forms. Below is a partial list of cultivated plants in the Brassica genus, along with their uses::
- white mustard (spice) (Brassica hirta aka Sinapsis alba)
- black mustard (spice) (Brassica nigra)
- rapeseed (Brassica napus)—grown for oil and animal feed
- canola oil (varieties of Brassica rapa and B. napus) — grown for oil and animal feed
- leaf mustard, brown mustard (Brassica juncea) —grown as a vegetable
- Ethiopian mustard, Ethiopian rape (Brassica carinata)—grown as a vegetable
- bok choy, rapini (aka brocoli rabe), turnip, napa cabbage (Brassica rapa)— grown as vegetables
- cabbage, kale, broccoli, brussels sprouts, cauliflower, kohlabi (Brassica oleoraca) — grown as vegetables
- rutabaga (Brassica napus) — grown as vegetable
As was the case with wheat, polyploidy is significant in the relationships of these crops. The crops come from three diploid species, (B. nigra, oleracea and rapa), and three polyploid (tetraploid) derivatives, B. carinate, juncea, and napus), produced from hybridization with pairs of the three diploids.
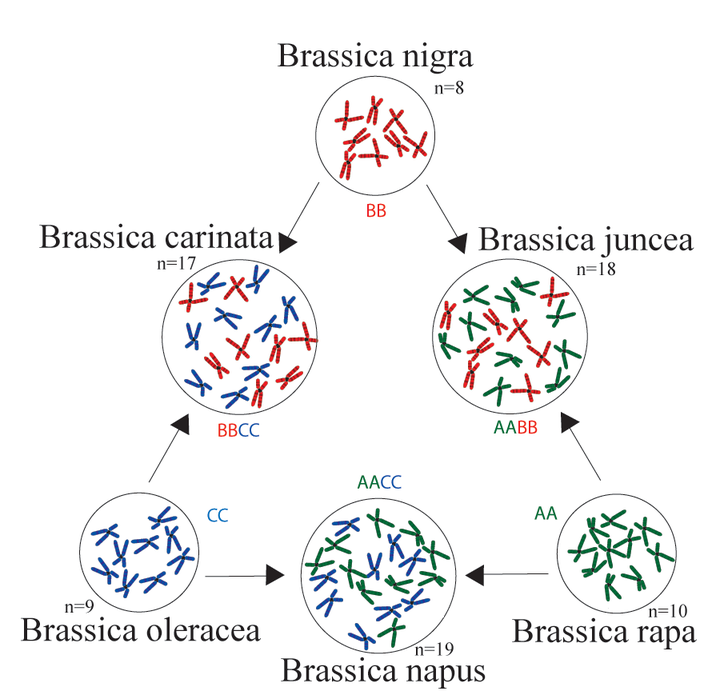
Another similarity to wheat, and to several other crops (e.g. sunflower) is that these species are ‘weedy’ (see Chapter 29), i.e. they thrive in ‘ruderal’ (disturbed) habitats. Hence the progenitors of most of these crops probably grew in the disturbed habitats that ancestral humans created by their behavior. Domestication followed from this. In fact, while black mustard is grown as a crop, it is also a significant weed in many parts of the world.
Unlike wheat, a polyploid group whose members all look very similar, the Brassica group shows remarkable morphological variation, especially the ones used as vegetables, where the edible part may be leaves (kale, collard greens, leaf mustard, Ethiopian mustard), roots (turnips, rutabagas), enlarged stems (kohlrabi), shortened stems with overlapping leaves (cabbage, see Chapter 10), lateral buds (brussel sprouts, see Chapter 10), and thickened inflorescence meristems (cauliflower, broccoli).
All of these crops were developed thousands of years ago by selection for plants that had favorable traits; i.e., ‘primitive’ agriculturalists noted favorable features and perpetuated plants that had these features. A challenge to these early genetic engineers is that most of these crops are biennials, meaning that some of the crop must be allowed to overwinter and in the second year transform into a flowering form from which the seeds are collected, i.e. these farmers needed to be aware that flowers and seeds are only available in the second summer after planting.
Another interesting feature of these plants is their chemistry. They produce a group of chemicals called glucosinolates, that, if cells are ruptured, are converted into chemicals called isothiocyanates. It is these chemicals that give members of this group (also closely related radish, horseradish and wasabi) a peppery and sometimes ‘hot’ taste. Glucosinolates release isothiocyanates as a result of the action of the enzyme myrosinase. This enzyme is sequestered separate from the glucosinolates, and only acts to release isothiocyanates when herbivory is occurring, crushing cells and breaking open cellular components, thereby allowing myrosinase to be in contact with glucosinolates. Although many humans find isothiocyanates desirable, many insects do not and the system acts as a chemical deterrent to herbivory (although, as is often the case, some insects specialize on mustards and are attracted by the chemistry). A comparable sequestering system operates in some plants (e.g. apple seeds) where cyanogenic glycosides are stored separate from enzymes that can cleave a sugar group to release cyanide.
Generating variability
Whether or not one is utilizing hybrids, crop development requires variability followed by selection of plants with favorable features. Open pollinated crops generate variability both through naturally occurring mutations and also by chance shuffling of genes in the sexual process. Although there had been suspicions on the existence of sex in plants and the roles of pollen and pollination, the observations of Rudolf Cameraius late in the 17th century revealed the role of pollen and made possible the generation of variability by the intentional crossing of two related species. Often these efforts were stimulated by the botanical explorations and the collection of ‘new’ plants, e.g. the production of commercial strawberries described above. This allowed closely related plants to be grown together and crossed. Early in the 20th century, workers discovered that they could generate variability with treatments that induce mutations: specific toxic chemicals or harsh radiation. And most recently, modern molecular techniques allow for the most precise modifications of plant characteristics through the introduction, or occasionally removal, of specific genes. Although the later process is the only one described as ‘genetic modification’, one should appreciate that crops have been genetically modified once they started to be cultivated. One should also appreciate that modern techniques are by far the most specific, meaning that the modifications are the most targeted, with the least amount of disruption of non-target genes and features (see Chapter 31).
Further Reading and Viewing
- “A Brief History of the Seed Industry” by SeedStory.
- “Date update: Methuselah gets six siblings” by The History blog. Date palm.
- “The extraordinary diversity of Brassica oleracea” by The Botanist in the Kitchen. Brassica crops.
- “Domestication, Genomics and the Future for Banana” by J. S. Heslop-Harrison and Trude Schwarzacher.
- “Domestication of the banana” by ProMusa.
- “George Beadle’s Other Hypothesis: One-Gene, One-Trait” by John Doebley
- “Strawberry: A Brief History” by David Trinklein.
- “Domestication of Plants in the Americas: Insights from Mendelian and Molecular Genetics” by Barbara Pickersgill.
- “Here’s what 9,000 years of breeding has done to corn, peaches, and other crops” by Brad Plumer.
- “Pirates of the Carob Bean” by The Botanist in the Kitchen.
- “Explaining Gluten” by Cooking Science Guy.
- “The Difference Between Waxy and Mealy Potatoes” by Cooking Science Guy.
- “The Unfortunate Sex Life of the Banana” by Matt Castle.
- “Vanilla Mania” by Bernd Brunner
Media Attributions
- VanGogh © Vincent Willem van Gogh is licensed under a Public Domain license
- Black emmer wheat © Robert Flogaus-Faust is licensed under a CC BY (Attribution) license
- Bread Wheat Evolution © Borb is licensed under a CC BY-SA (Attribution ShareAlike) license
- Strawberry watercolor © Deborah Griscom Passmore is licensed under a Public Domain license
- Navel Oranges © Edward H. Mitchell is licensed under a Public Domain license
- Banana © Chris 73 is licensed under a CC BY-SA (Attribution ShareAlike) license
- Teosinte and modern corn © National Science Foundation is licensed under a Public Domain license
- Brassica oleracea var. gongylodes © mud is licensed under a CC BY-SA (Attribution ShareAlike) license
- Triangle of u simple © adenosine is licensed under a CC BY-SA (Attribution ShareAlike) license

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}