
Chapter 13: Sex and reproduction in non-seed plants
Almost all plants are capable of reproduction without sex. Most commonly this happens as a consequence of the plant being severed into pieces and these pieces being able to regenerate the parts that were lost. Additionally, some species have developed pieces that are particularly prone to be broken off, e.g., the gemmae (singular of gemma) of some mosses and liverworts. It is relatively common for flowering plants to produce asexual propagules in the places where seeds normally develop, producing seed-like structures without the normal process of sex. Although reproduction without sex is common in plants and for some species it represents their sole means of reproduction, most plants rely on sexual reproduction, and two innovations, the seed and the flower, have been particularly significant to the evolution of plants.
With respect to sex, the plant kingdom illustrates wonderful patterns of unity and diversification. On the one hand, all plant life shows a fundamentally similar pattern of sex, on the other hand, plants illustrate remarkable diversification of this common theme.
TOPICS
- Alternation of generations
- Mosses, liverworts and hornworts
- Ferns
- Horsetails
- Clubmosses
All plants exhibit alternation of generation s, they produce two types of multicellular organisms: one diploid and derived from the development of a zygote, and one haploid and derived from the development of a haploid cell called a spore. ‘Development’ involves the proliferation of cells, the differentiation of cells and the formation specific structures with specific roles, i.e.. morphogenesis.
All plants exhibit alternation of generations and alternate between haploid and diploid organisms. To complete the sexual cycle the haploid organism must produce gametes that unite to form the zygote. That is, among all the haploid cells that are produced as a result of cell divisions of a haploid spore, some of the cells are endowed with special capabilities that allow them to encounter and interact with another gamete to form a zygote. Similarly, of all the diploid cells derived from the zygote, some, called a ‘spore mother cells’, are endowed with the ability to undergo meiosis to produce haploid cells, spores, that grow into haploid individuals. Note that meiosis does NOT produce gametes, rather it produces spores that are dispersed and develop into haploid plants. The diploid plant that produces spore mother cells and spores is called a ‘sporophyte’. The gamete-producing organism, which is haploid, is called a gametophyte. Thus, there is an ‘alternation of generations’ with a haploid form alternating with a diploid form (Figure 1). Often one of the two forms is challenging to appreciate, largely because they are often challenging to see — they are often small, short-lived, and may not live a separate existence from the previous generation. That is, the haploid and diploid forms may not spatially distinct from each other; they may be temporarily, or permanently, a part of their parent!
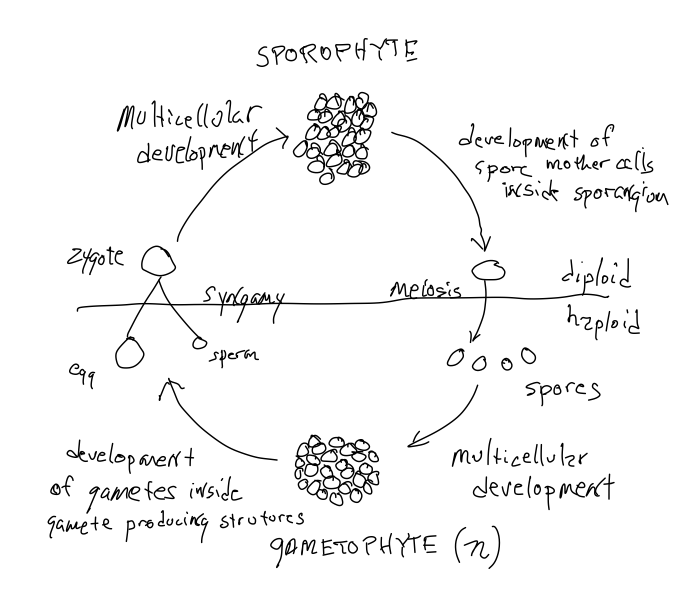
Like those of familiar animals, the gametes of all plants are of two types, a larger, immobile egg and a smaller sperm that must in some way be mobile in order to encounter the egg. The gametophytes of plants may be bisexual (hermaphrodites), producing both egg and sperm, or unisexual, i.e., there are separate male and female gametophytes. In some plant groups, including all of those discussed in this chapter, the sperm is mobile because they possess flagella. Sperm swim away from the male gametophyte that produced it and are chemically attracted to the structure producing eggs. Because plants are terrestrial organisms, the water required for a flagellated sperm to swim to the egg but water is not always present. We will see in the next chapter that some plants have circumvented this problem by having male gametophytes become mobile, not the sperm they produce.
As in animals, the female gamete, the egg, does not move. It is retained in the organism that produces it. In the case of plants this is a haploid gametophyte (not a diploid individual as in mammals). And in some of the plants discussed in this chapter the zygote formed by the fertilization of an egg grows out of the archegonium (the structure in which an egg is produced) and produces a sporophyte attached to the gametophyte. Alternatively, the entire (female) gametophyte of some plant groups is embedded in the diploid plant (the sporophyte) that produced the spore that developed into the female gametophyte. That is, in some plant groups, the spores produced by sporophytes are not dispersed; instead the spore germinates where it is produced and grows to produce a gametophyte plant on or inside a sporophyte plant. Mobile male gametophytes and sedentary female gametophytes are features of a very few of the non-seed plants discussed in this chapter but are ubiquitous in seed plants, discussed in the next two chapters. In this chapter we consider the more readily observable and understandable alternation of generations that is found in plants without seeds.
Mosses Liverworts and Hornworts

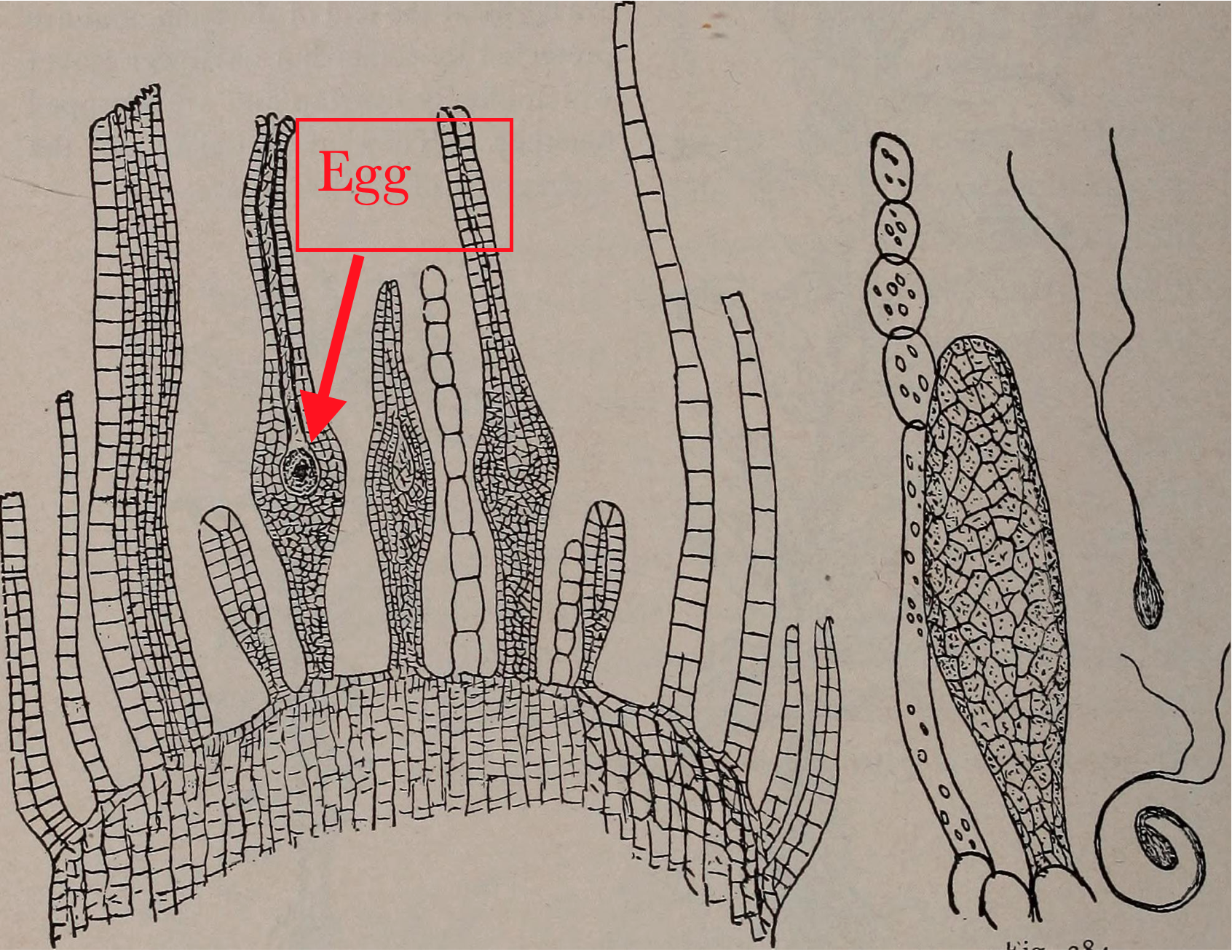
In mosses, liverworts and hornworts (the three groups of plants lacking vascular tissue) it is the gametophyte plant that lives the longest and is the most visible. It produces a form capable of gathering materials for growth, primarily through photosynthesis (Fig. 2). The most common form, found in all mosses and many liverworts, is an elongate axis bearing flaps of tissue that increase the photosynthetic area. At some point, often once a year, the gametophyte produces gamete producing structures that are called archegonia (singular = archegonium) if they produce eggs and antheridia (singular = antheridium) if they produce sperm. These structures are typically produced in the midst of modified ‘leaves’ at ends of stems/branches in mosses or, in some liverworts, they occur on the underside of umbrella/mushroom-shaped structures that extend upwards from the main body of liverworts.. The antheridia (Fig. 3) are roughly spherical containers in which cells differentiate into flagellated sperm and are released when the antheridia break open. The archegonia are flask shaped structures with a single egg near the base and an elongate neck that develops a canal through which sperm can swim in order to fertilize the egg. The resulting zygote develops inside the archegonium, producing a sporophyte that eventually extends typically one to six cm beyond the archegonium and exists as an appendage of the gametophyte, never liv ing an existence independent from it. Although the sporophyte sometimes is green and capable of photosynthesis for part of its life, its structure, a simple stalk with no ‘leaves’ to increase surface area, is not particularly suited for photosynthesis. Rather the structure is suited for the dispersal of spores; the stalk usually elevates the capsule, which is the site where spore mother cells develop, undergo meiosis, and produce spores.
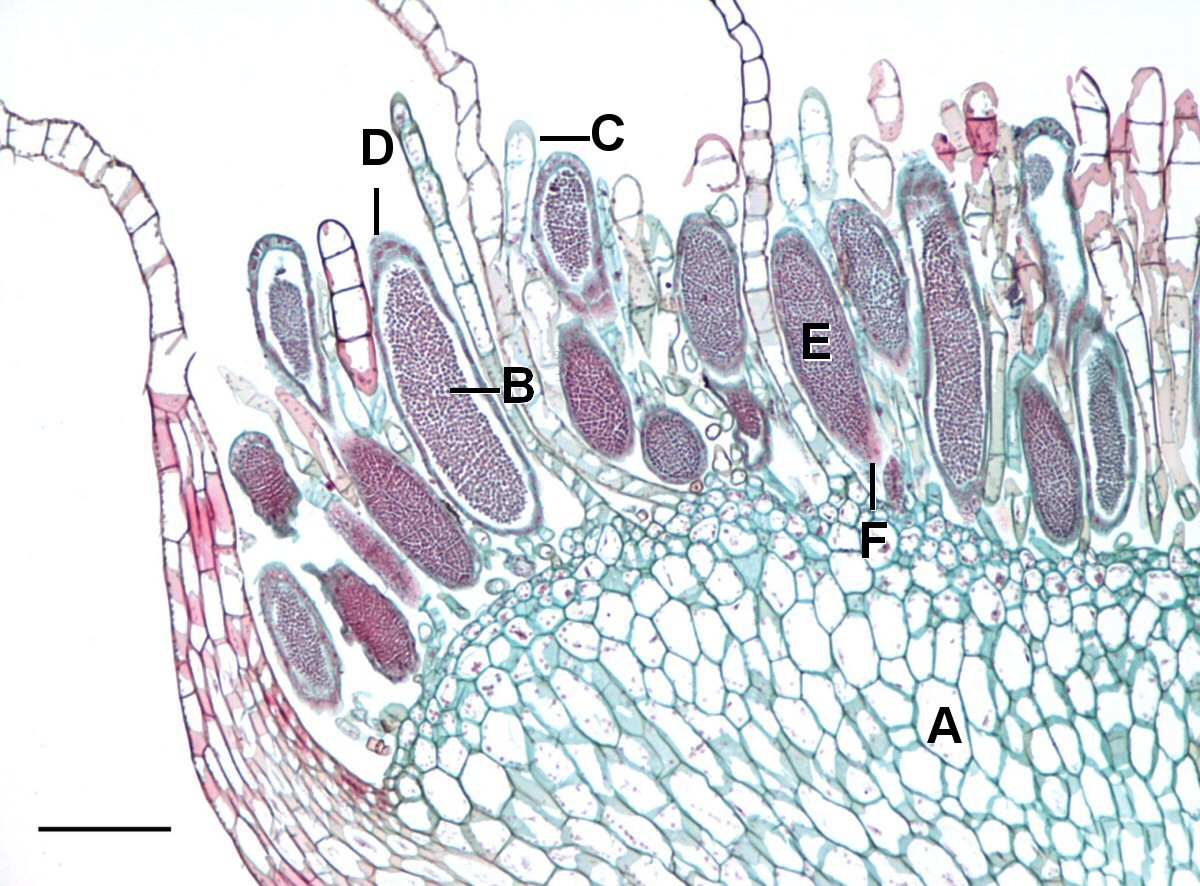
If ‘spatial separation’ is used rigorously to define an organism then these sporophytes would not be considered separate organisms, they are simply a part of the gametophyte (Fig. 5). But, in light of the sexual cycle and pattern of alternation of generations, it is helpful to consider them to be separate organisms. This is supported by fact that they have a different number of chromosomes than the gametophyte that they grow out of.

In all other plants besides mosses, liverworts and hornworts it is the spore producing plant that lives the longest and is the most visible; it is the form that we see and recognize as a plant. The sporophyte still begins its development growing out of the archegonium where the egg was produced and was fertilized, but the sporophyte’s growth is such that it becomes completely independent of the gametophyte that it emerges from and eventually has a completely autonomous existence. Thus, when we see ferns, horsetails, club-mosses, and seed plants, what we are observing is a diploid plant that produces spores. In all these groups the gametophyte is small and elusive but the basic life cycle is the same as in all plants: an alternation of generations between a gametophyte and a sporophyte. Seeds and seed plants are discussed in the next chapter, below are considered aspects of the sexual cycle, in particular features of the less commonly seen gametophytes, that are produced by seedless vascular plants.
Ferns
Most ferns have a small, photosynthetic gametophyte that usually is less than 1 cm across and one cell thick, i.e., a sheet. It lacks a stem axis and is often ‘heart-shaped’ (Fig. 6-8 and 11). It is generally attached to a substrate via rhizoids (filaments of non-photosynthetic cells).

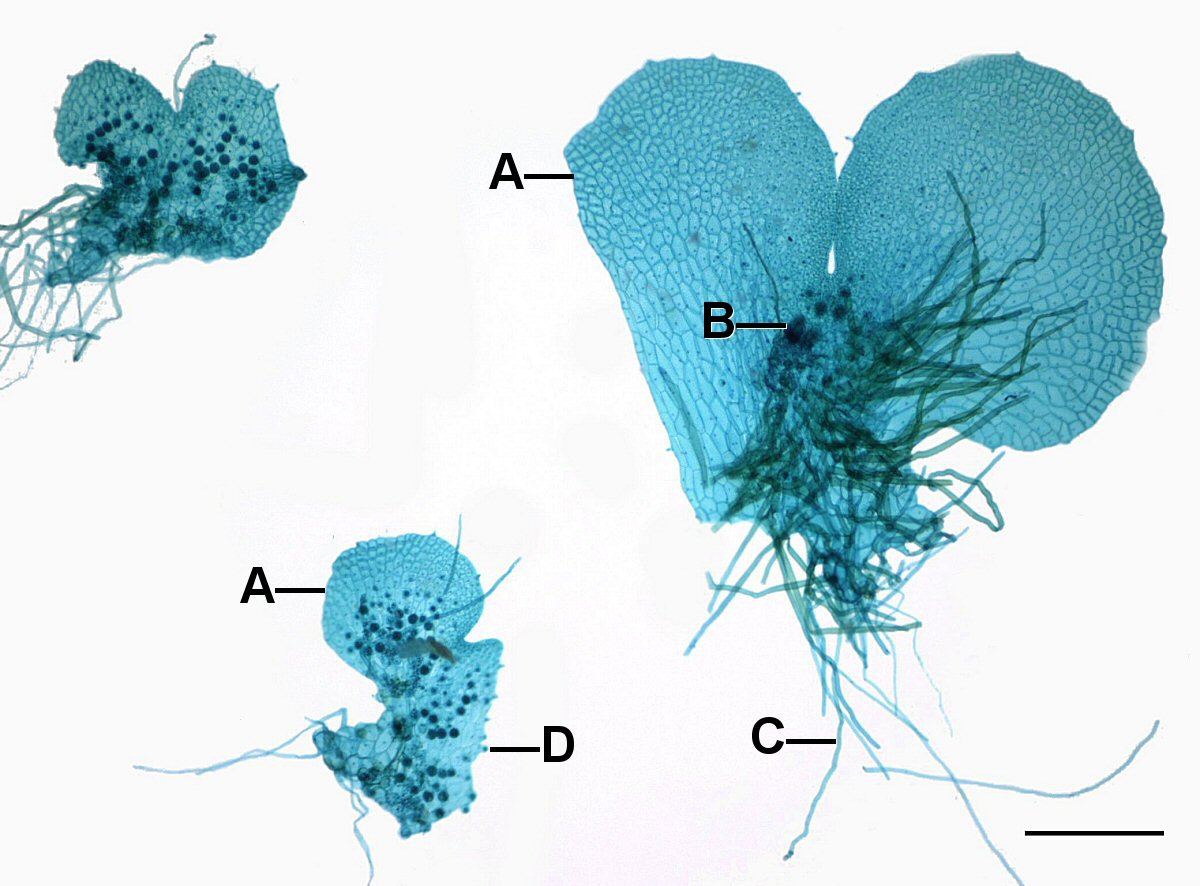
As was the case in the mosses, the fern gametophytes produce structures where the egg and the sperm are produced as a result of cells dividing in a particular pattern to produce archegonia (Fig. 7-9) and antheridia. It is important to note that gametes are not produced by meiosis because all the cells of the gametophyte are haploid already.

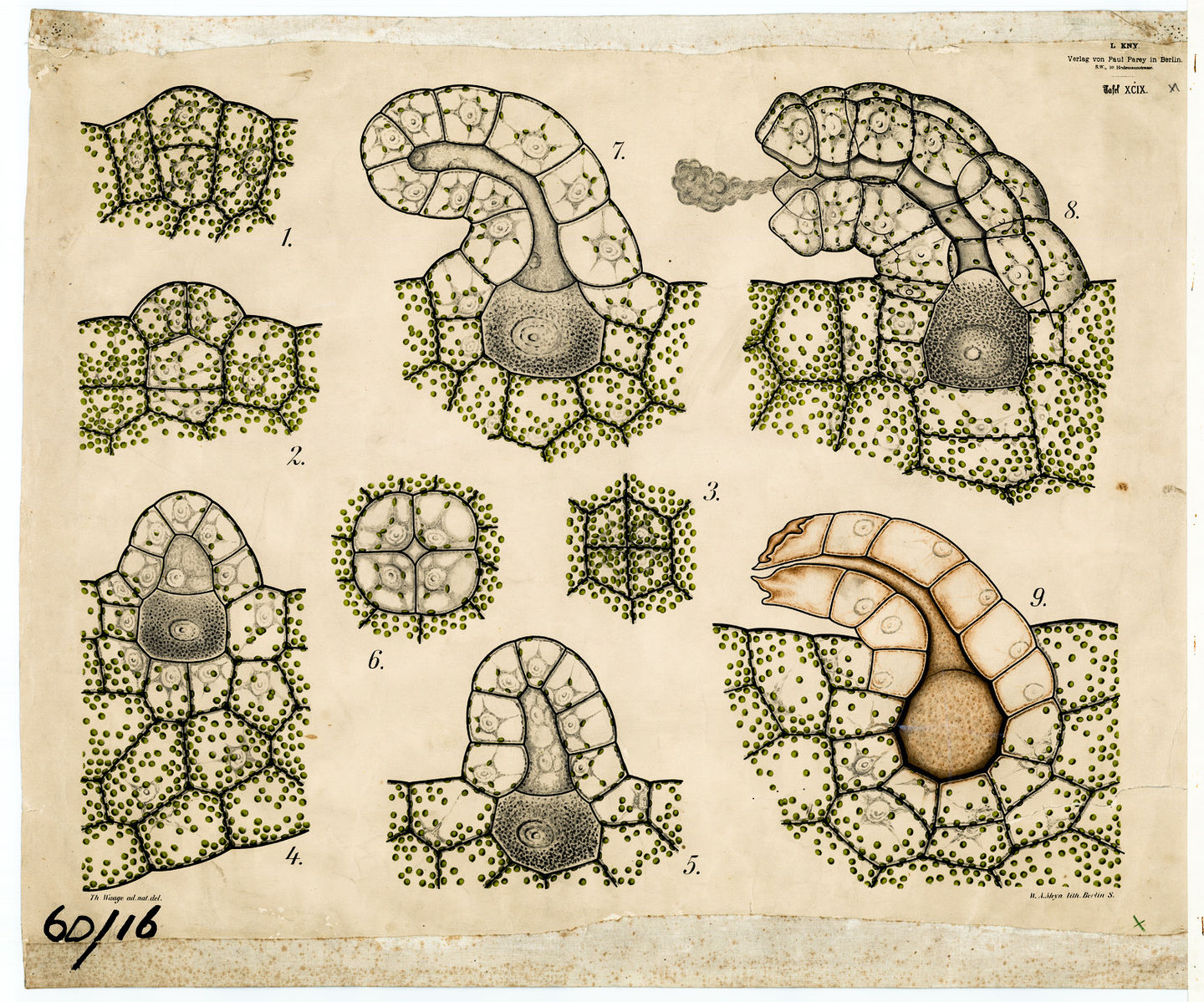
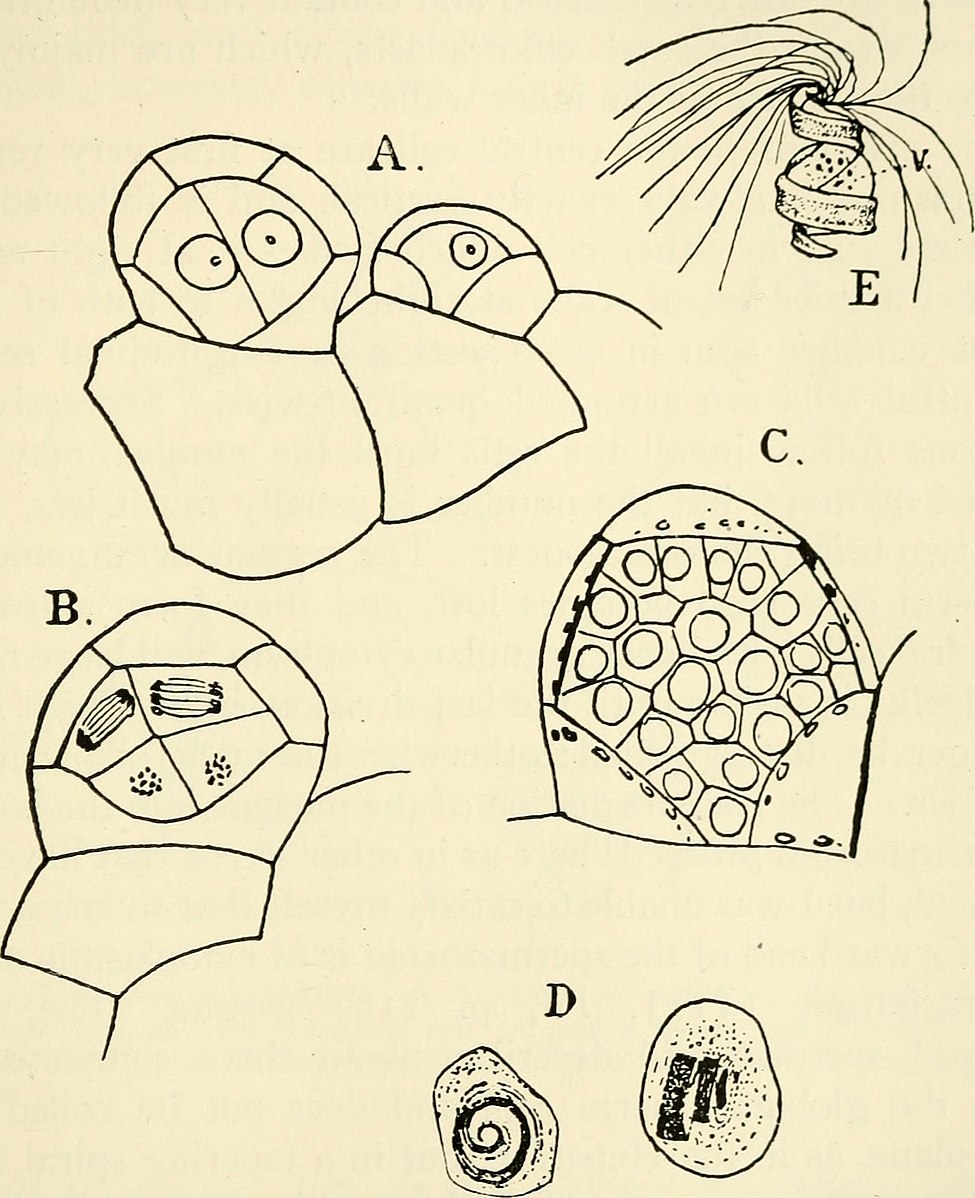
Fern gametophytes are generally have flask shaped female structures (archegonia) located in the notch between the lobes and globular male structures (antheridia) located on the on the lobes. While most fern gametophytes are hermaphroditic, some are unisexual and for some their sexual expression depends on environmental conditions. All the cells of the gametophyte are haploid but it produces a cell, the egg, with special developmental abilities. The antheridia release sperm that have flagella which allow them to swim to the archegonia, where the eggs are located, and swim down a narrow canal to fuse with the egg cell at its base. The zygote develops into a sporophyte, producing stems and roots. The stems produce leaves which shade the gametophyte and it soon dies (Fig. 9)

The sporophyte continues to grow to produce the fern that we recognize. It has the same structure as most plants: a root-shoot axis with leaves produced by the shoot. Most of the ferns in this area have stems (rhizomes) that are below ground and relatively short. What we see are the leaves emerging above ground from this rhizome. At some point this diploid organism produces structures termed sporangia, inside of which are spore mother cells that undergo meiosis to produce a group of four (a tetrad) of haploid spores that are released to the environment. When these germinate, they grow into haploid gametophytes and the process is repeated. Generally, the sporangia are produced in clusters called ‘fruit dots’ that are located on the underside of leaves (see wood fern). Other ferns have entire portions of their leaves that are obviously different and where spores are produced. A few ferns in this area are dimorphic (see sensitive fern), producing two types of leaves, some that are green and photosynthetic and which never produce spores and other leaves that are non-photosynthetic and produce abundant spores, while being nourished by the photosynthetic part of plant.
While this is the general pattern for ferns, there is some variation, one example of which is the water fern Marsilea, an aquatic fern that has several interesting features (see the information sheet on Marsilea). As is the case in a number of ferns, spores are produced on a specialized leaf that is very different looking from normal photosynthetic leaves. Whereas the normal leaves are green and shaped like clover leaves, the spore bearing leaves are packaged into a seed-like structure, hard on the outside and capable of being dried out and revitalized (germinated) when re-wetted. At this time the ‘fruiting’ (i.e., spore bearing) leaf emerges into the water, looking very little like a leaf: it is without chlorophyll, very small and gelatinous. It produces spores in clusters. There are two types of spores, male spores called microspores and female spores called megaspores, each in separate sporangia. The technical name for plants that produce two types of spores is heterosporous. In contrast, most ferns are homosporous, producing only one type of spore that generally produces hermaphroditic (bisexual) gametophytes (described above) ; a few homosporous forms produce unisexual gametophytes, both male and female, but both coming from identical looking spore s.
The two types of spores of Marsilea are readily distinguished by size. The megaspores are around 1 mm in length and germinate to produce egg-producing, female gametophytes. The microspores are only ~ 70 um in length and produce sperm-producing male gametophytes. Without any increase in size and without emerging from the microspore, the male spores germinate to produce a ‘plant’ with ~35 cells, 32 of them are spermatazoids, multiflagellated corkscrew shaped sperm cells, roughly 10 um in size, that are released from the male gametophyte when the microspore wall is broken. The spermatazoids are chemically attracted to the female gametophyte. The female gametophyte is substantially larger than the male gametophyte but it still is small and, like the male gametophyte, exhibits endosporic development, meaning that its development occurs within the spore case of the megaspore, with only the very short neck extending from it. It produces a single archegonium with a single egg that the sperm swims to and fertilizes, forming a zygote. While the new sporophyte plant seemingly sprouts from the female spore, it actually is coming from a female gametophyte that is growing inside the spore case.
Further Reading
“Marsilea: Habitat, External Features and Reproduction” on Biology Discussion.com
Another interesting fern is the Appalachian bristle fern, which is only known from the gametophyte form. Apparently, it has been reproducing asexually for millions of years! There are several other species of ferns known only as gametophytes.
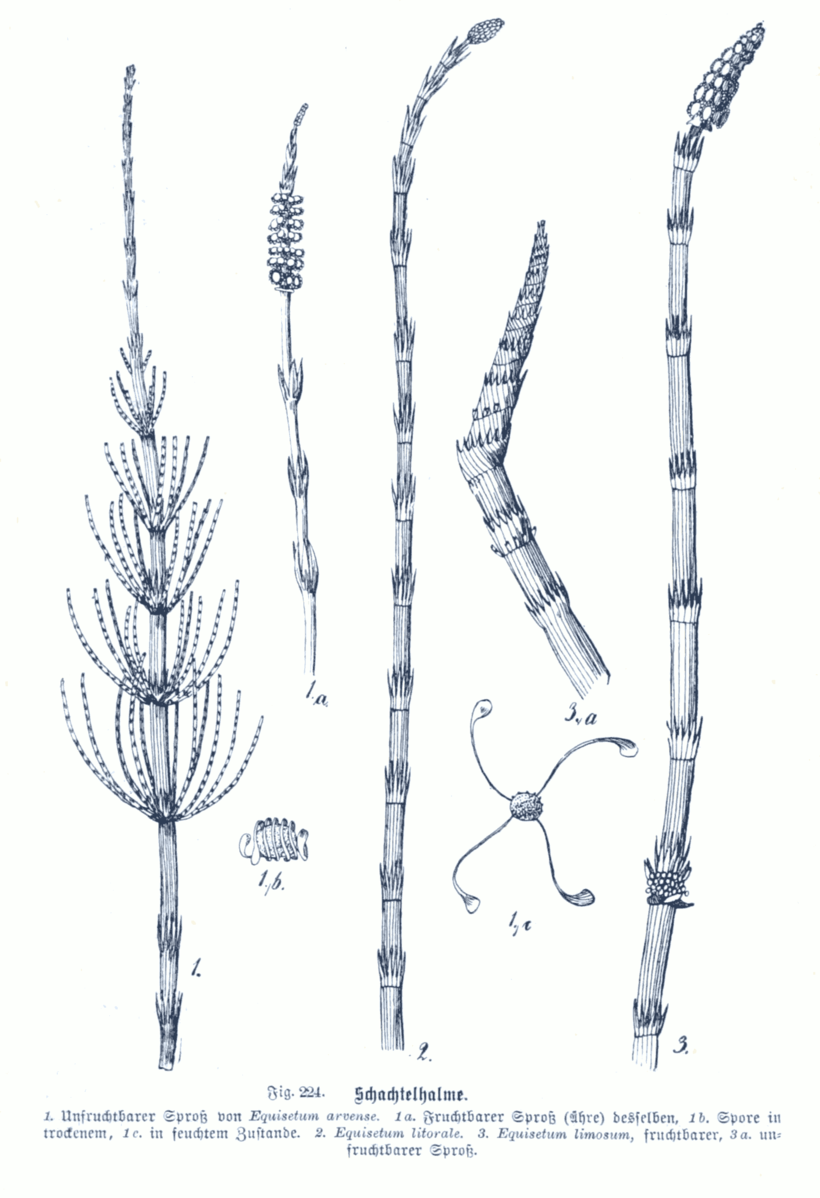
Horsetails
The basic pattern found in ferns, with a dominant sporophyte generation and a diminutive gametophyte generation, is found in the horsetails, a group of vascular plants that originated in the Paleozoic and produced a number of tree forms that were significant in producing extensive deposits that became coal and oil. There only remains one genus of horsetails and there are less than 20 species worldwide. All are herbaceous with perennial rhizomes that send up vertical branches that have a very distinctive pattern of growth with photosynthetic stems, very small scale-like leaves, and whorled branches or no branches. Spores are produced in a terminal cone-shaped structure, which is a cluster of specialized, spore bearing leaves. Spores are dispersed by the wind but their movement and release from the sporangium may be aided by structures called elators, strap-like appendages on the spore that move in response to the absorption and loss of water. Germination of the spore produces a small (~ 1-3 cm) photosynthetic gametophyte that looks like a pin cushion. Very short ‘stems’ are present with appendages that increase photosynthetic area. Antheridia and archegonia are usually both produced from the same gametophyte, although it may be unisexual for a period of time. Sperm are multi – flagellated and need to swim to reach the egg. Fertilization results in a zygote that develops into a diploid sporophyte that soon overgrows the gametophyte that it emerges from, producing roots and both horizontal stems (rhizomes) and vertical stems.
Clubmosses, Spikemosses and Quillworts
The se three groups are thought to be closely related and are grouped together as ‘Lycopods’ or ‘Lycophytes’, a group that also includes a number of fossil forms, including tree-like forms that were very important at the end of the Paleozoic era. The group has 1200 species and is considerably more diverse than the horsetails but much less diverse than the ferns (12,000 species). The sporophytes of extant clubmosses, spike mosses and quillworts are all herbaceous perennials. They generally spread extensively with above-ground and below-ground stems (tropical members are usually epiphytes). Although ancient members of the group exhibited woody growth, none of the species alive today do.
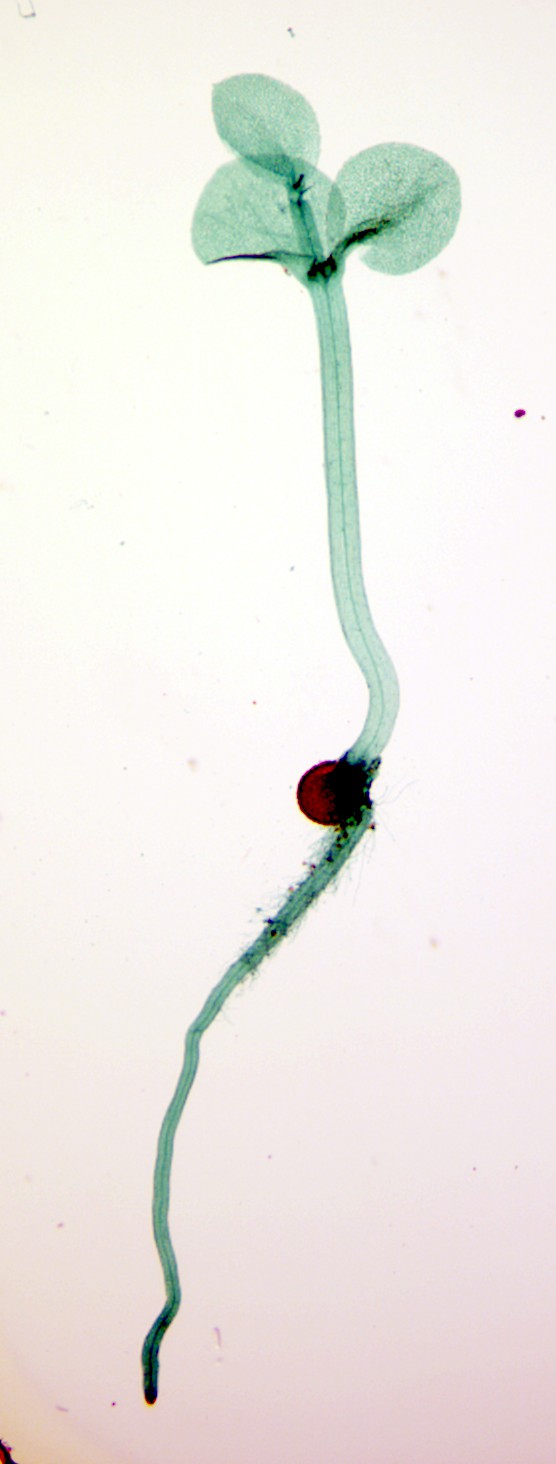
Clubmosses (Fig. 13) are unique in having gametophytes that are subterranean and non-photosynthetic, surviving as parasites by feeding off of fungi. These gametophytes live much longer than most gametophytes of vascular plants, some over 15 years. Spikemosses and quillworts are heterosporous and, like the aquatic fern Marsilea, the gametophytes develop endosporically; living off the material that was provisioned in the spore by the sporophyte plant. The male gametophyte is very short-lived and has little stored material (the microspore is small), but the female gametophyte is considerably bigger and lives for months on material present in the spore. Early growth of the sporophyte, out of the female gametophyte that is present in a megaspore, looks like a germinating seed (Fig. 14) and we will see in the next chapter that these are analogies — it is not thought that seed plants developed from the lycopods. Another feature of some spikemosses is that the ‘female’ spores, the ones that develop into female gametophytes, sometimes develop ON the sporophyte plant, a nother feature that is repeated in seed plants.

Further Reading and Viewing
- “Information about preparations for the Science Rendezvous (Bring a Fern Baby Home for Mother’s Day) event” by Markham. Fern life cycle (nice images).
- “Bryophyte Ecology” by Janice M. Glime. Bryophyte ecology, great site, more than just ecology.
- “Australian Bryophytes – Bryophytes” by Heino Lepp. Bryophytes, hornworts, liverworts.
- “Australian Bryophytes – What is a liverwort?” by Heino Lepp. What is a liverwort
- “Living together and living apart: the sexual lives of bryophytes” by David Haig. Bryophyte sex.
Media Attributions
- Antheridia © Jon Houseman is licensed under a CC BY-SA (Attribution ShareAlike) license
- Antheridium © George Francis Atkinson is licensed under a Public Domain license
- Fern Gametophytes L © Jon Houseman is licensed under a CC BY-SA (Attribution ShareAlike) license
- fern model 1
- Archegonia © Paul K is licensed under a CC BY (Attribution) license
- The structure and development of mosses and ferns © Douglas Houghton Campbell is licensed under a Public Domain license
- Onoclea sensibilis © Vlmastra is licensed under a CC BY (Attribution) license
- Root and shoot © Curtis Clark is licensed under a CC BY-SA (Attribution ShareAlike) license

{kind=link}
_(14760367676).jpg){kind=link}
{kind=link}
.jpg){kind=link}
_(1918)_(14784806482).jpg){kind=link}
{kind=link}
{kind=link}