
Part I: An Introduction to Paleoanthropology
5. What is a Hominin
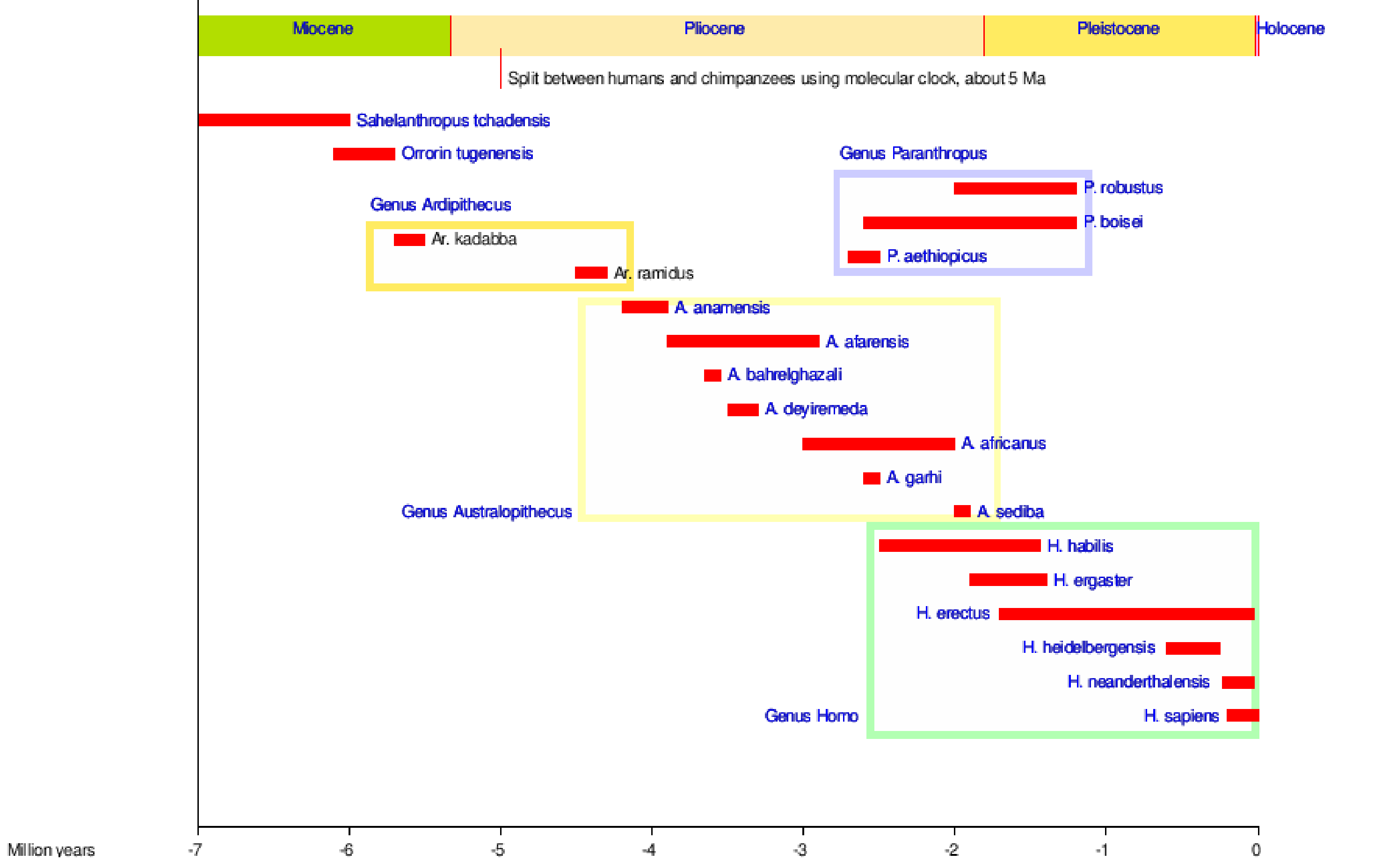
We are hominins, as are all those bipedal apes that came before us. Figure 5.1 shows most of the hominin species through time, some of which we are descended from and some that are side branches in our tribal tree!
ANATOMICAL TERMS
In order to adequately understand a discussion of hominin evolution and appreciate changes over time, some basic anatomical information is necessary. It is also necessary in order to distinguish primitive or ape-like skeletal characteristics from those that are derived, i.e. those that arose later in time.
Any discussion of anatomy assumes that one is speaking of a body in anatomical position, i.e. facing forward if standing or supine (face up) if lying down, with palms forward or up (see Figure 5.2—5.4). When referencing particular structures or regions of the body, we make use of the following terms:
- Superior—closer to the top of the head.
- Inferior—closer to the soles (or plantar surfaces) of the feet.
- Medial—closer to the midline of the body.
- Lateral—closer to the far right or left of the body, relative to the midline.
The following two terms are used in reference to the limbs:
- Proximal—closer to the base of a limb.
- Distal—closer to the end of a limb.
You may hear your professor use the above terms when describing aspects of human or fossil specimen anatomy.
Note: If you are thinking about a career in paleoanthropology, get as much training in anatomy as possible, even while an undergraduate. Anthropology and biology departments may offer courses in human anatomy, human osteology, forensic anthropology, and the like. Finds like “Lucy” (Australopithecus afarensis, see Chapter 11) come along once in a lifetime. Most discoveries consist of little pieces of fossilized bone or teeth and thus knowledge of human anatomy is crucial for recognizing possible landmarks on the remains.
HUMAN GROSS SKELETAL ANATOMY
The terms gross anatomy and macroscopic anatomy refer to the study of structures that are visible to the naked eye. In this section, we will consider whole bones versus the individual parts of bones and only those bones that are external, as those are most relevant in a discussion of bipedalism and human evolution. While the anatomy of the lower limb takes precedence in a discussion of bipedalism, other parts of the body also changed over time. The anatomy of the skull is particularly important because the skulls and faces of hominin species changed over time and geographic space.
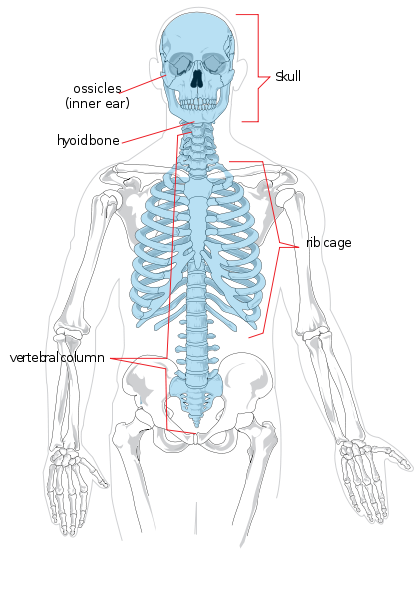
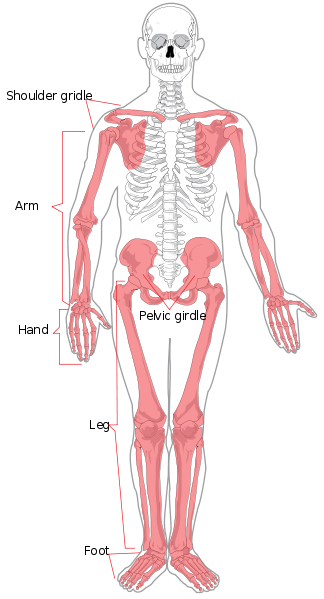
We will begin with regional anatomy. The axial skeleton consists of the head and torso. Regionally it is divided into the skull, thorax, and spinal column. The spinal column is divided into the seven cervical (neck), twelve thoracic (thorax/chest), five lumbar (lower back), five sacral (also known as sacrum), and four coccygeal (also known as coccyx or tailbone, they may also number three or five) vertebrae. The limbs and shoulder and hip regions make up the appendicular skeleton. The upper limb is also known as the arm or forelimb. It is divided into the arm (upper arm), forearm, wrist, and hand and fingers. The lower limb is also known as the leg or hindlimb and is made up of the thigh, leg (equivalent of forearm), ankle, and foot and toes.
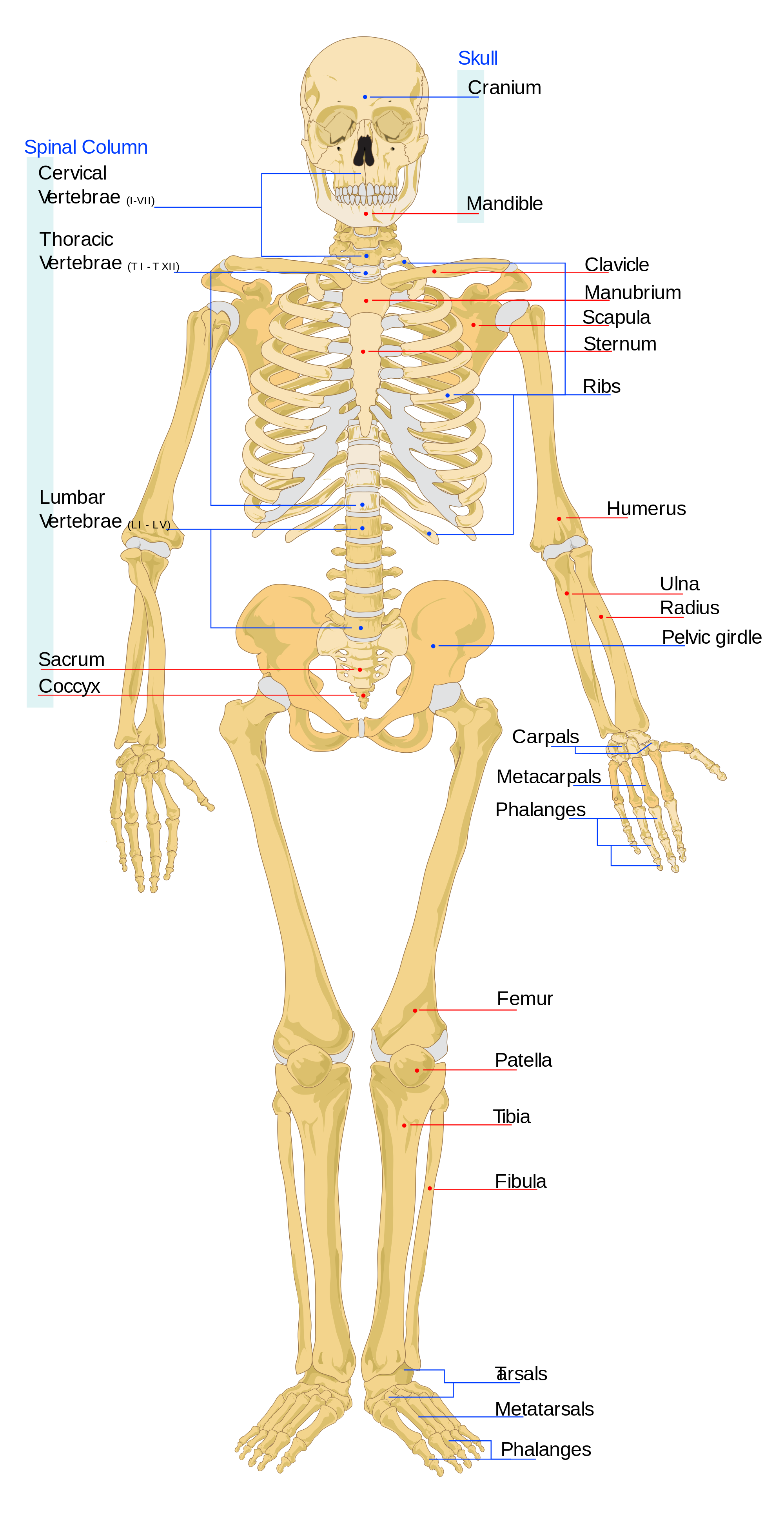
Individual bones of the axial and appendicular skeletons are labeled in Figure 5.4. They will not be discussed here.
THE EVOLUTION OF BIPEDALISM

There are a variety of theories as to how bipedalism evolved and why it proved to be so successful for early hominins. One early idea suggested that by standing up, our ancestors would have been able to see above the grass and thus avoid predation. Baboons and patas monkeys provided living models for hypothesizing the environmental stresses early hominins might have faced on the open plains of Africa. While they likely traveled through open areas, we now know that the earliest hominins were exploiting forest resources, as evidenced by their thinner molar enamel, relative to later hominins. There were also theories that involved the freeing up of the hands to make and use tools and for carrying resources to a safe place or home base. C. Owen Lovejoy believes that bipedalism allowed males to provision mates with resources (Lovejoy, 1981). Those males with the most advanced bipedal capabilities would have had an increased chance of mating and possibly offspring survival, and thus bipedalism would have spread throughout the population. While Lovejoy makes a good case for how a trait could be favored in a population, it is not clear why females would have needed to be provisioned unless their offspring had already lost their ability to hang on with their feet, and hence became a burden to foraging. We now know that ardipiths, while bipedal on the ground, had a divergent hallux, so that young animals could likely have hung on to their mothers in trees like modern primates. Although we cannot know for sure whether they were covered with hair, I can only speculate that when on the ground, young infants may have hung by clasping the fur on their mother’s ventrum (anterior aspect of the trunk) while the mother supported their bottom. As the baby matured, it may have clasped its hands around the mother’s neck or chest and hung on to her fur with its feet and later ridden “piggy-back” like modern quadrupedal monkeys and apes. Eventually, it would have walked beside her from place to place. However, if resources had become extremely scarce, bipedal males may have ventured out onto dangerous ground for resources with which to provision their mates. Another theory that sees males as being the impetus for bipedalism suggests that males may have been more terrestrial and females more arboreal, i.e. a case of niche partitioning, like gorillas and the mandrill and drill monkeys, where males forage on the ground and females and young spend more time in the trees. Other theories also suggest that bipedalism was a response to the changing nature of the resource base. For example, Meave Leakey and Kevin Hunt (a theory known as Hunt’s Postural Feeding Hypothesis, Hunt 1996) believe that the ability to stand on two legs for long periods of time would have facilitated picking fruit from the terminal branches of low, scrubby trees in the increasingly open habitats of East Africa. While the aforementioned theories are not mutually exclusive and there was likely a synergistic effect that resulted from our ancestors’ changing locomotor capabilities, a highly plausible model suggests that it was our ability to break out of the “ape habitat” that facilitated our evolutionary success.
The following items will help us to better understand this great “step” in our ancestry:
- Apes are descended from an arboreal climber from the early Miocene of Africa.
- The ancestor of the African great apes possessed a suspensory hanging adaptation and thus had an upright trunk that was wide and shallow; mobile shoulder and wrist joints; long arms relative to leg length; long, curved hand and foot bones; and an opposable big toe (hallux). The next section explains how that thoracic morphology facilitated our ancestors’ bipedalism.
- The common ancestor of chimps and hominins was likely a semi-terrestrial quadruped that was adapted to climbing, feeding, and sleeping in trees, as well as moving and foraging on the ground.
- The late Miocene of Africa was marked by climatic change that spurred floral and hence faunal changes. Equatorial Africa was cooler and increasingly drier than in earlier times. The Indian subcontinent continued to move north, resulting in the uplifting of the Himalayan mountain chain that produced a rain shadow, i.e. moisture-laden clouds that would have previously drifted down into Africa now lost their moisture on the mountains. Drying winds and cooler temperatures resulted in forest reduction and fragmentation in Africa, splitting and isolating resident faunal populations.
- The majority of African ape species went extinct due to loss of habitat.
- Bipedalism is an efficient means of locomotion for covering distance on fairly level ground.
- A more vertical posture reduces the surface area exposed to solar radiation in a more open environment. It also raises a large percentage of the body away from the hot ground, where it is exposed to cooling breezes.
- The ancestors of the hominins adapted to the changing environment by becoming bipedal on the ground. Over time they developed a more efficient heat exchange system for their bodies (sweating) and brains (large venous sinuses for rapid blood movement) and lost much of their body hair.
- Based upon early hominin teeth, they were generalists like chimps, likely getting the majority of their carbohydrates and fats from fruit, protein from young leaves, and possibly fat and protein from animal matter, e.g. social insects (chimps and gorillas eat a lot of ants and termites) and animals caught opportunistically. No early hominins exhibit the same degree of canine size or sharpness as chimp and gorilla males. Their canines stay sharp via a honing (sharpening) action with the first lower premolar, termed a sectorial premolar due to its unicuspid morphology. The combination of the action and morphology of the two teeth is termed a “honing complex”. Males’ canines are exceptionally large and, in combination with their high degree of sexual dimorphism, are used to compete with other males for access to females. Of course, they are also useful for predator defense. Thus, if the common ancestor of chimps, gorillas, and hominins possessed a premolar honing complex, as seems likely, the early hominin fossil evidence suggests that they were losing their fighting teeth. In addition, fossil hominins do not exhibit the same degree of sexual dimorphism as seen in chimps and especially gorillas. Later chapters contain more information on hominin socioecology.
Most apes went extinct as their habitats dwindled and they competed for limited resources. However, with an efficient means of locomotion to move between forest patches when resources became depleted, hominins could continue to exploit those resources to which they were adapted. They also likely evolved new capabilities for exploiting newly encountered food items as they moved through and between ecozones. Loss of habitat and resources often leads to local extinctions. By expanding their home ranges and dietary niches, hominins survived while the majority of their close relatives did not.
BIPEDAL ANATOMY
See Figure 5.4 for individual bones.
The majority of bipedal characteristics involve the hip (or pelvic girdle) and lower limb. However, as will be seen below, certain skull and trunk characteristics are also adaptations for bipedal locomotion. In addition, we have inherited many aspects of our upper bodies from our ape ancestors and those will all be discussed in the following sections.
SKULL
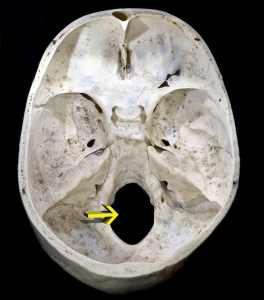
The skull consists of the bones of the braincase and face and the mandible (lower jaw). The foramen magnum is the hole in the occipital bone situated in the base of our skulls (see Figure 5.6). It is where our spinal cord exits the cranial vault. In hominins, the foramen magnum is positioned more anteriorly than in the other apes because our head sits on top of our vertebral column. Thus while the earliest hominins had very ape-like faces, the position of the foramen magnum shows that they were bipeds.
VERTEBRAE
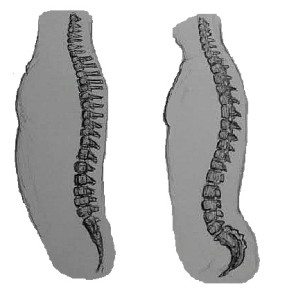
Ape spines are not as flexible as monkeys’ spines, giving us better upper body support since we are more upright than most other primates. Our vertebrae increase in size and robusticity from top to bottom so that our lumbar vertebrae are very large; they sit on the fused vertebrae of the sacrum, which is firmly attached to the hip bones. The sacrum is large and broad and curves inward (as does the coccyx) to help support the organs. Thus our spinal column is a strong supporting structure for the upper body. We hominins have two larger curves in our backs relative to the other apes, the cervical curve and the lumbar curve. The fact that our heads are more upright than nonhuman apes means that the cervical vertebrae must form a more concave curve, i.e. the superior aspect of our neck is arched back relative to theirs (see Figure 5.8). The more pronounced lumbar curve forms when we stand up and begin toddling about. The joints between the lumbar vertebrae are easily strained and it is thus important to maintain strong back and abdominal muscles throughout life, to aid in the stability of the region.
THORAX
The thorax consists of the sternum, ribs, and thoracic vertebrae. The ape thorax is adapted to climbing and swinging in trees. It is wide (right to left) and shallow (front to back) relative to quadrupedal monkeys, which have a narrow and deep thorax like those of dogs. While the morphology of the thorax was originally adapted to arboreal climbing, the upright trunk also allowed for bipedal locomotion. In addition, the shallow nature of the thorax brings the center of gravity closer to the vertebral column for better weight concentration and transfer. Apes are better bipeds than most nonhuman primates. They can walk bipedally for short to moderate distances, depending on the species, but it is not efficient and they cannot maintain it for very long.
SHOULDER GIRDLE
The shoulder girdle consists of the clavicle, scapula, and humerus. The shoulder joint of extant nonhuman apes and early hominins is and was angled upward, demonstrating the arboreal ancestry of those hominins and, in combination with their long, curved fingers, it suggested that they could ascend and climb about in trees. Our clavicles stabilize our shoulder joints for swinging and hanging. The triangular shape of the scapula (shoulder blade) is more equilateral than that of a quadrupedal monkey’s, which is more elongated. Our scapulae are more mobile than those of a typical quadruped and the articular surface of the scapula, i.e. where the round head of the humerus articulates (makes contact), is shallow and allows us to rotate our arms at the shoulder. This suspensory hanging complex of clavicle, scapula, and humerus morphology (also elbow and wrist adaptations—see below), along with the muscles and connective tissue involved, allows us to climb, hang, and swing by our arms.
UPPER LIMB
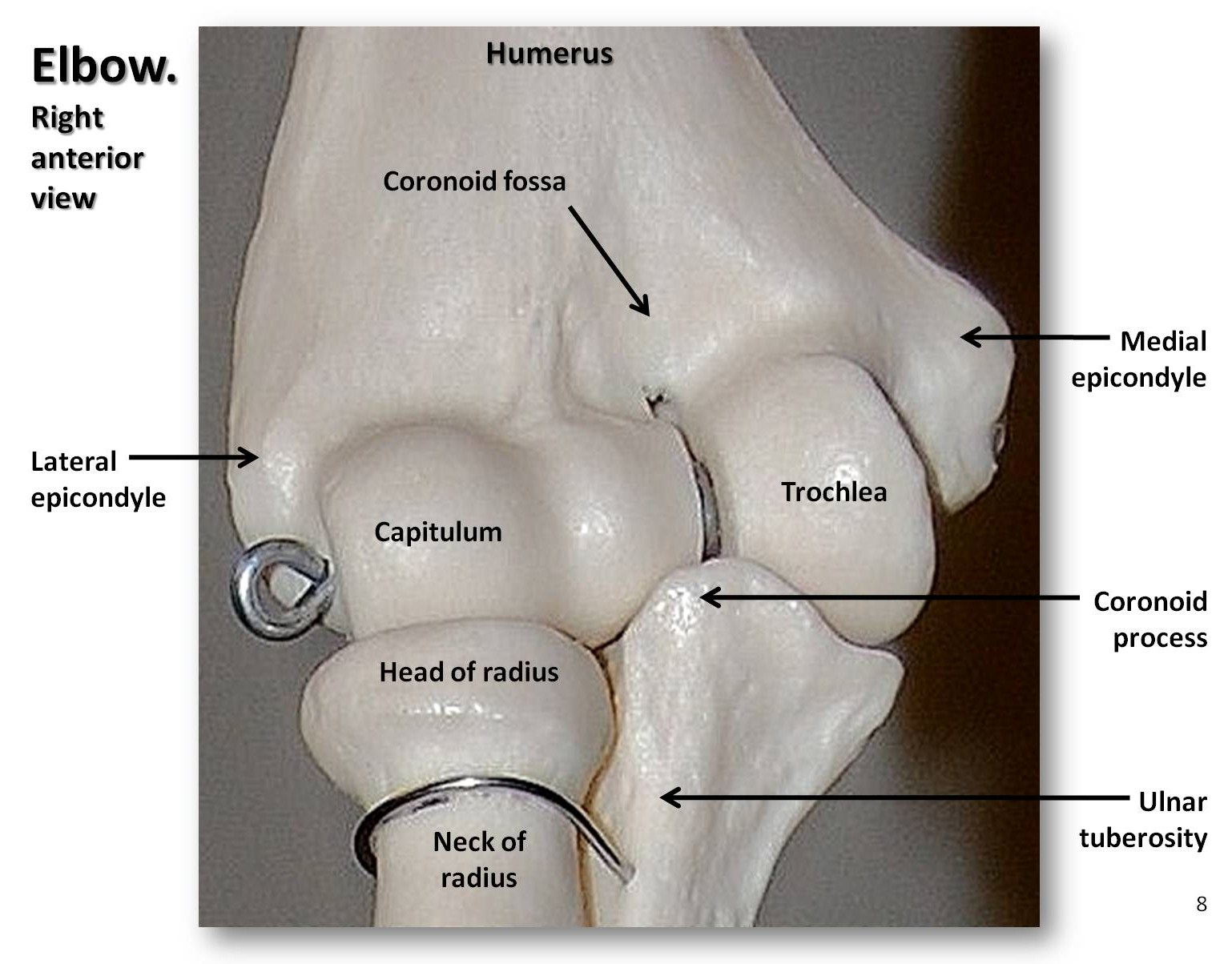
The upper limb consists of the humerus of the arm, the radius and ulna of the forearm, the eight carpal bones of the wrist, the five metacarpals of the body of the hand, and the phalanges of the digits (three per finger and two per thumb, or pollex). As mentioned, the head of the ape humerus is round, resulting in a very mobile shoulder joint. We can fully extend our arms at the elbow in order to hang or swing, whereas quadrupeds cannot. In addition, we can supinate and pronate our forearm, i.e. move our hand palm up or palm down. This movement is possible because the head of the radius is a concave disk that rotates on a ball-like structure termed the capitulum on the distal humerus (see Figure 5.9). Our wrist joints are very flexible, allowing us to rotate and twist our hands in a variety of ways. Early hominins had short legs, long arms, and curved fingers. Over time, hominin legs lengthened so that their intermembral index (IMI) became reduced. The intermembral index is the ratio of armlength to leglength, calculated by the following equation:
[latex]IMI =(\frac{arm length}{leg length})100[/latex]
If an animal has long legs and short arms (like us), they have a low IMI and vice versa; if fore- and hindlimbs are approximately equal in length, such as in terrestrial quadrupeds, the IMI will be close to 100.
HIP OR PELVIC GIRDLE
Our shoulders are somewhat analogous to our butts! This is because muscles originating from outside the limbs are crossing the joints to insert upon and move the limbs. Because we are bipeds, it is not as apparent as it is in quadrupeds, whose fore- and hindlimbs move similarly. However, if you compare the humerus and femur and the muscles that cross the respective joints (deltoids and gluteal muscles, respectively), you will definitely see similarities.
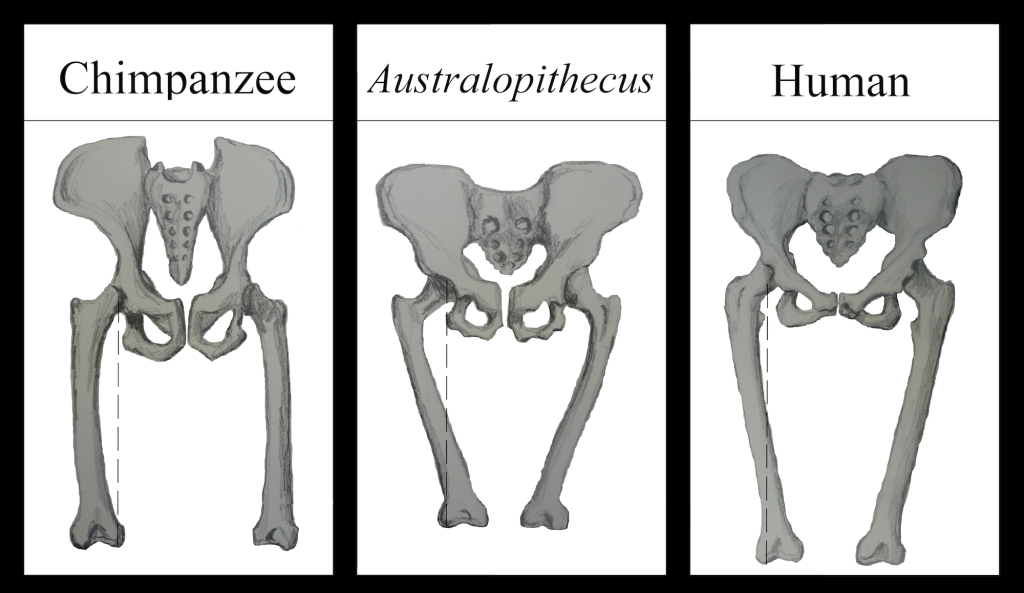
Our pelvis is very unique and interesting. It has changed significantly from an ape pelvis (see Figure 5.10). The pelvis is made up of three bones: the two lateral bones, termed innominates or os coxae, and the sacrum. Collectively, they form a basin-like structure that holds our internal organs while providing support for our upper bodies. Each innominate consists of three bones that fuse during development: the ilium, ischium, and pubis (see Figure 5.13). They meet at the hip joint. Hominin innominates became shorter and broader, so that the ilium wraps around laterally from an earlier, more posterior position. This changed the orientation and action of our hip muscles, allowing for our striding gait and the ability to balance our weight on one fully extended leg while the other leg is in the swing phase. A portion of the gluteus maximus muscle inserts behind the hip joint in hominins (versus lateral in chimps), and thus instead of abducting the femur (moving it out laterally, as when doing jumping jacks), it changed to a powerful hip extensor (backward motion) for running.
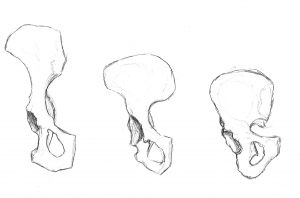
Figures 5.11 and 5.12 illustrate the landmarks that are adaptations to bipedalism. The iliac crest is long and curved, as the bone wraps around laterally. The iliac blade is short but expanded horizontally. The iliac blades are buttressed or reinforced to handle the stress of our upper body weight. The thick section of bone, termed the iliac pillar, can be seen running from the iliac crest (at the iliac tubercle) down behind the hip joint. The articular area, termed the acetabulum, is large and deep, providing a stable socket for the ball-like head of the femur. The joints of the pelvis are very strong and relatively immobile (compared to the shoulder joint). The sacroiliac joint (between the ilium and the sacrum) is large and more posterior and proximal to the hip joint than in quadrupedal apes. Together with the strong pubic symphysis (anterior joint where the pubis portions of the two innominates meet), these characteristics make for a very stable supporting structure. 
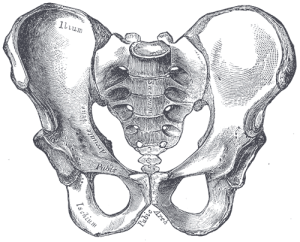
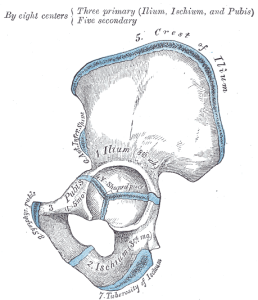
“Plan of ossification of the hip bone” by Henry Vandyke Carter is in the public domain.
LOWER LIMB
The lower limb consists of the femur of the thigh, the tibia and fibula of the leg, seven tarsal bones of the ankle, five metatarsals of the body of the foot, and phalanges of the digits (three per toe and two per big toe or hallux). The head (proximal ball-like structure) of the hominin femur is large. The femur angles medially (inward) from hip to knee, so that our upper body weight is transferred down through our hip joints to our knees. This is termed the carrying or bicondylar angle. The knees of quadrupedal apes are directly below the hip joint, so there is more strain on the knee joints when they walk bipedally (see Figure 5.13). We have two bulbous structures on the bottom of each femur, termed condyles. The innermost condyle, the medial condyle, has lengthened in hominins so that the femur sits on top of the flat tibial surface. If this were not the case, the medial condyle would not make contact with the tibial plateau, due to the bicondylar angle. Weight is transferred through our innominate, hip joint, and lateral condyle of the femur (see Figure 5.13, "Human"). Unlike apes’ knees that are chronically flexed, our knees are capable of full extension; each locks into place when the other leg is in swing phase, giving us a stable supporting leg. Each gluteus medius muscle alternately supports the opposite side of the torso and pelvis, so that it does not slump on the unsupported side.
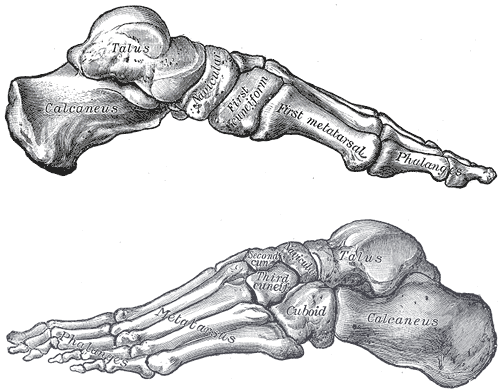
Our feet (see Figure 5.14) have changed dramatically from a mobile, grasping structure to a rigid, supporting one. The tarsal bones of the human ankle are large and robust for support. The joint between the distal tibia and fibula is fairly immobile, so that the two bones are firmly lashed together. Together, they articulate with the talus (most superior tarsal bone) in a hinge joint. We have lost much of the mobility of an ape foot and thus have become less agile in climbing over time. The calcaneus or heel bone is very large and robust and, along with the ball of the foot (distal end of the first metatarsal) and the area below the baby toe (fifth metatarsophalangeal joint), forms a tripod structure. Our feet have three arches for support, shock absorption, and forward propulsion; they are the medial and lateral longitudinal arches and the transverse arch. Hominin toes became shorter and less curved over time. While shorter, the hallux became more robust and lost its degree of divergence and opposability, by coming into alignment with the lateral four toes, i.e. digits II through V.
HOMININ EVOLUTIONARY TRENDS
Just as there are evolutionary trends that characterize primates, there are also trends that characterize the hominins, i.e. traits that developed over time and occur to varying degrees in the various hominin species (see Table 5.1). While all of the morphological changes involved with bipedalism are hominin trends, there are also relevant characteristics in other parts of the body.
| Table 5.1 HOMININ EVOLUTIONARY TRENDS |
| Bipedalism and related morphological adaptations. |
| Encephalization and corresponding prolonged juvenile dependency. |
| Loss of craniofacial robusticity and muscle attachments. |
| Reduced prognathism. |
| Reduction in size of dentition. |
| Molarization of premolars. |
| Increased manual dexterity. |
| Elongated legs and increased stature. |
| Increased reliance on culture, technology, and language. |
| Increased reliance on meat in the diet. |
Most notable, our ancestors and their relatives became increasingly more intelligent. Our brains have increased in size more than four-fold, from a more chimp-sized brain (<400 cc) in the earliest hominins to a mean of ~1400 cc. This likely occurred in response to environmental stresses as well as competition with other hominins for resources. Skull size and shape changed in response to encephalization, i.e. increasing brain size.
Brains are very costly organs and researchers believe that in order for brain size to have increased, there would have had to have been a corresponding decrease in some other costly organ system. It is hypothesized that a higher quality diet allowed the hominin gut to shrink and, in turn, the brain to expand. Marked encephalization in the hominin lineage began with the first members of our own genus: Homo. While there is some evidence that earlier species (e.g. australopiths) manufactured tools, there is solid evidence that early Homo did, and the archaeological record suggests an increasing reliance on meat in their diet. While the dimensions of the thorax remained wide and shallow, the shape of the rib cage changed from a more conical, ape-like thorax that could accommodate a large gut, to our more barrel shape. As our brains became bigger, our ancestors gave birth to increasingly altricial (helpless) young that continued their development outside of the womb. Our offspring require protection and care for a much longer period than those of the other great apes, i.e. we have a prolonged juvenile dependency period. At least by the time of the earliest members of the erectus grade at 1.8 mya (see Chapter 27 and subsequent chapters), it is thought that infant requirements necessitated assistance from mates and/or relatives.
Many of the early hominins had pronounced, forward-oriented jaws, termed prognathism (pro = forward; gnath = jaw). Over time, hominins became more flat-faced, or orthognathic. While extant African apes retain primitive prognathism and the shearing/honing dental complex, hominins lost those pronounced canines, as well as the gaps in the corresponding tooth rows—termed canine diastema (singular) or diastemata (plural)—that allow apes to close their jaws. The hominin first lower/mandibular premolar developed into the characteristic bicuspid (two-cusped) morphology over time, as opposed to the apes’ unicuspid sectorial premolar. The loss of the honing complex may have been in response to reduced competition for females as a result of male-female pair-bonding. Once the jaws no longer locked into place, our ancestors were free to grind their food, aiding in its digestion.
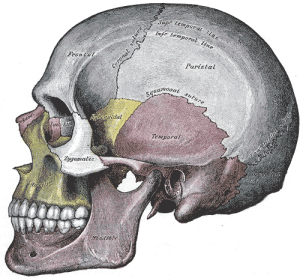
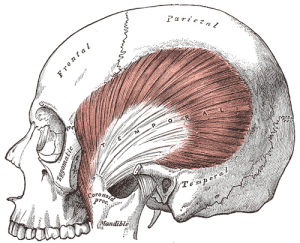
The size of jaw and neck muscle attachment sites on the skull became reduced in the hominin lineage over time, along with a reduction in the size of the teeth and craniofacial robusticity. The action of the powerful temporalis muscle (a muscle of mastication) changed from primarily acting on the front of the jaw, allowing apes to clamp their jaws powerfully shut during fighting, to acting on the molar region for grinding food (see Figure 5.15). The origin of the temporalis muscle moved over time from the midline of the top of the skull to a more inferior position on the lateral aspect of the frontal and parietal bones (see Figure 5.15), due to the reduction of the sagittal crest and decrease in temporalis power in hominins.
Leglength increased as our hominin ancestors came to rely less on an arboreal environment and became primarily terrestrial. The pronounced increase in leglength and stature seen in the erectus grade (see Chapter 27), relative to past species, allowed them to cover vast distances efficiently and they moved from Africa to Asia in a relatively short period of time.
Hominin fingers became shorter and lost their curvature over time. By the time of the australopiths, hands had become more dexterous. There is evidence that Australopithecus africanus possessed a “power” thumb, giving them increased abilities for holding objects in one hand while manipulating or working them with the other hand. This was necessary for our ancestors to have made and efficiently used tools. The first metacarpal was adapted for such activity, and the intrinsic thumb muscles (within the hand versus extending from the forearm into the hand) were well-developed. There is some controversial evidence that by the time of the later robust australopiths, or paranthropines, expanded fingertips had appeared. Thus they may also have possessed the enlarged tactile pads and increased sensitivity and vascularization that allows for modern humans’ fine degree of manual dexterity.
Finally there is the cultural side of things. We are like no other species that has ever lived. We are unique in our ability to practice, produce, and pass on culture. Our brains are predisposed to the acquisition of language and culture. Thus, the final evolutionary trend seen in the hominins is an increased reliance on culture and language over time.
HOMININ GROUPS
The hominins can be divided into five groups that will be covered in subsequent chapters, based on shared characteristics and/or phylogenetic affinity:
- Earliest bipeds: Orrorin and possibly Sahelanthropus.
- Forest-adapted bipeds with relatively thin molar enamel: ardipiths.
- Bipeds that exploited a more open and drier niche with thick molar enamel: gracile australopiths, such as Australopithecus anamensis and afarensis.
- Descendants of gracile australopiths with heavy masticatory apparatus, adapted to exploit tough or hard foods when necessary: paranthropines, also known as robust australopiths.
- Hominins that retained the gracile masticatory apparatus of their australopith ancestors and exhibited a trend for encephalization and increasingly complex culture: Homo species.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}