
Part III: Pliocene Epoch
15. Australopithecus africanus
Australopithecus africanus (3–2 mya)
(“southern ape” / Africa)

SITES
Taung, Makapansgat, Sterkfontein, Drimolen, and Gladysvale, South Africa
PEOPLE
Raymond Dart, Robert Broom, and C. K. Brain
INTRODUCTION
Australopithecus africanus was the first fossil hominin discovered in Africa. In 1924, Raymond Dart (see his biographical sketch this chapter) identified the face, mandible, and endocast as being that of a juvenile bipedal ape (see Figure 15.1). Eugène Dubois’s discovery of the Javanese Homo erectus fossils in 1891 refuted the reigning belief that “we got smart before we stood up.” Once Dart’s claims were accepted, the world realized the extent to which that idea was false. The small-brained Au. africanus showed that early hominins were bipedal apes as opposed to quadrupedal humans.
Unlike East African material that can be dated using a variety of techniques, primarily due to past volcanic activity, South African fossils and sites are much more difficult to date. Most dates can only be represented as ranges; hence the date for the earliest Au. africanus specimens falls between 3 and 2 mya.
PHYLOGENY
Au. africanus is considered to be a gracile australopith by some and a robust australopith by others. Traditionally, the species was favored as the immediate ancestor of the Homo lineage, specifically of Homo habilis. However, some researchers have always believed that Au. afarensis was the common ancestor of both Au. africanus and the Homo lineage, suggesting a cladistic event had occurred at Au. afarensis. This schema has gained popularity in recent years. However, with the new evidence being put forth for Au. sediba, it seems that the Au. africanus → Homo scenario was closer to the truth in that Au. africanus and Au. sediba are undoubtedly related and Au. sediba shares many characteristics with genus Homo. While there is some support for Au. africanus as ancestral to the more derived robust forms, that still leaves unresolved those characteristics shared between Au. aethiopicus and both P. boisei and Au. afarensis (see Au. aethiopicus, Chapter 17).
DISCOVERY AND GEOGRAPHIC RANGE
Au. africanus is known only from sites in South Africa (see map showing the major fossil sites in Chapter 11, Figure 11.3). Material from more than 200 individuals has been collected over more than 80 years. Most of the fossils came from caves, some of which were discovered during mining and blasting activities. Those caves formed via underground water activity. Fossilization was facilitated by water dripping on bones and calcifying, just as stalagmites form. Over millennia, many of the caves filled in with mineralized deposits and as the ground surfaces eroded, the underlying deposits were exposed and mined and then later excavated for fossils.
BIOGRAPHICAL SKETCH: RAYMOND DART (1893–1988) and the “TAUNG CHILD”
Raymond Dart was the first to recognize a fossil hominin in Africa. While his claim to have discovered a human ancestor was not initially accepted by the scientific community, he was vindicated when Robert Broom began finding similar material at other South African sites. The work of Dart and Broom played a pivotal role in directing attention to Africa as the birthplace of humanity, as Charles Darwin had earlier predicted.
Raymond Dart was a renowned Australian anatomist who was teaching at the University of Witwatersrand in South Africa at the time of the discovery. I have read several accounts over the years of the fateful day when the fossil material landed in Dart’s hands. The Taung material was brought to him by either quarry workers or a colleague while he was dressing for the wedding of his daughter or friend, depending on the source you read. He is said to have attempted to use his wife’s knitting needles to extract the fossils from the surrounding matrix. I am not sure why we need to know what he was doing when he got his hands on the goods. We certainly do not know what most paleoanthropologists were doing when they were presented with fossil finds. However, it conjures up amusing possibilities in my mind!
The fossilized remains came from the Taung Quarry in the process of blasting for lime. When presented with the material, Dart established that it was the face, mandible, and endocast (fossilized interior of the cranial vault) of a juvenile hominin. He based this on the anterior position of the foramen magnum and aspects of brain morphology reflected on the interior of the skull vault. Dart named his find Australopithecus africanus, meaning “southern ape of Africa,” and the specimen became known as the “Taung Child.”

The Taung child was two to three years old when it died. We now know that the caves of South Africa were formed by underground water activity and hominins that dropped or were dragged into those subterranean caves became fossilized during the process of speleothem formation. Over millennia, the caves filled with mineral deposits, and via erosion, the mineralized contents and underground cavities surfaced. Many fossils were likely destroyed during quarrying activities, but we are lucky that many were preserved.
Raymond Dart is also renowned for his “Killer Ape” theory and the osteodontokeratic (bone-tooth-antler) culture. Dart believed that animal long bones and carnivore mandibles that were found with australopith remains had been used as weapons to fight and kill one another. We now know that was not the case. They were most likely opportunistically hunting small prey and scavenging larger kills, and they were prey for larger animals.
Raymond Dart is credited with the 1924 discovery and naming of Au. africanus. His now famous “Taung Child” came from the Taung quarry site. The two- to three-year-old juvenile is represented by its face, skull fragments, and mandible, and an endocast of its brain. Dart also worked at the site of Makapansgat. His contemporary, Robert Broom (see biographical sketch below), worked at the caves at Sterkfontein (see Figure 15.4), where he discovered a complete, female cranium known as “Mrs. Ples”, along with other Au. africanus material. Broom also worked at the sites of Kromdraai and Swartkrans; the latter is where he discovered the first paranthropine, Paranthropus robustus. C. K. Brain, a famous taphonomist, also worked at Sterkfontein. He discounted Dart’s views of the australopiths as “killer apes”. He believed that bones either dropped into caves as part of large cats’ prey or were dragged in by rodents for gnawing. Some individuals are thought to have accidentally been trapped in underground caverns. Some speculate that those individuals that show no evidence of having been preyed upon, due to their degree of completeness (e.g. Sts 573 from Sterkfontein and the Au. sediba party from the Malapa site), surely became trapped. The six Au. sediba individuals are thought to have possibly been attracted to the cave by water. Drimolen, a more recently discovered site, has yielded an almost complete cranium as well as material from approximately 80 individuals. A fifth site associated with the species is Gladysvale.
BIOGRAPHICAL SKETCH: ROBERT BROOM (1866–1951)

Robert Broom was a Scottish medical doctor and paleontologist who subsequently made a name for himself as a paleoanthropologist, even though the term did not exist at the time. He taught geology and zoology at a South African college until he was let go for his controversial beliefs in evolution that were contrary to the religious teachings at the college. Subsequent to his termination, his finances went downhill until Raymond Dart’s influence secured him a position at the Transvaal Museum. While his specialty was mammalian-like reptiles, he became increasingly involved with fossil hominins. Together, Dart and Broom made paleoanthropological history, discovering the first and second species of African hominins, respectively. Broom worked at the Au. africanus sites of Sterkfontein and the P. robustus sites of Swartkrans and Kromdraai. All of those sites are now contained within the Cradle of Humankind World Heritage Site. He was the first to discover and name Paranthropus robustus, and his work with Au. africanus helped to support Dart’s claims to have discovered a bipedal ape and human ancestor. Broom’s most famous Au. africanus find was “Mrs. Ples” (possibly a male), which he originally named Plesianthropus transvaalensis or “primitive human” of the Transvaal.
If accounts of Robert Broom are to be believed, he was a colorful character. He is said to have worn semi-formal attire while excavating and when media were present, he conveniently happened upon important discoveries. Supposedly, one of his team buried an artifact that was already labeled with a catalog number. Oh, to have been on the scene and witnessed what must have been an embarrassing situation! Not to mention that, according to numerous online sources, he thought nothing of stripping naked when it became “Africa Hot”! (I picked up that term from a couple of comedy movies but it is now in the Urban Dictionary.)
It is interesting that Broom did not believe in Darwinian evolution but rather, what we would now call Intelligent Design (Wikipedia contributors 2015h).

PHYSICAL CHARACTERISTICS
Au. africanus was more derived than Au. afarensis. This is not surprising considering that they lived at least one million years later, as well as the trend within the hominin lineage to become more encephalized and manually dexterous over time. Au. africanus were habitual bipeds with all of the corresponding lower limb adaptations. They also retained climbing characteristics, such as upward-oriented shoulder joints; long arms relative to legs; and long, curved hand and finger bones. However, in general their hands were more human-like than those of Au. afarensis.
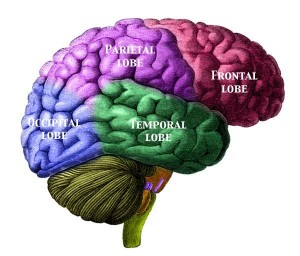
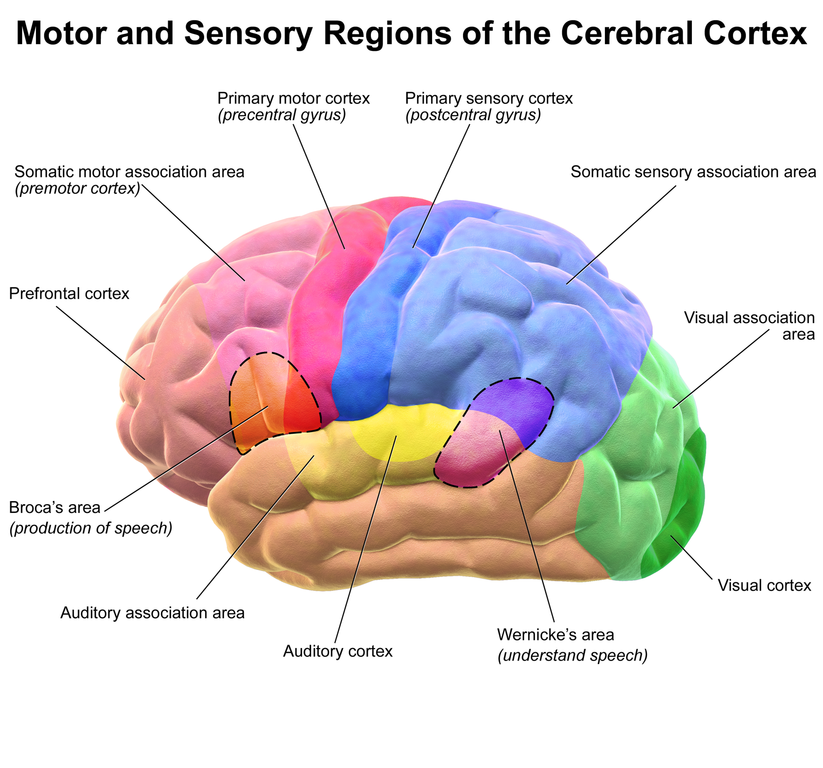
While the brain was small relative to later species, Au. africanus was not only more encephalized than past species, with a cranial capacity of 450 cc (range = 424–508 cc), but also possessed an enlarged cerebral cortex in the frontal and parietal regions (see Figure 15.5). Their encephalization quotient (EQ) was 2.7. The quotient is a method for comparing brain size among species. Anything greater than 1.0 means that the brain is larger than would be expected based on body size (FYI: our EQ is ~7.6). Broca’s area is an area of the left lateral frontal cortex that is involved with the production of language (see Figure15.6). It is present in all Old World monkeys and apes, but it is enlarged in Au. africanus relative to previous species. These are all important developments, in that they herald the appearance of more complex thought processing and likely communication skills. There is debate over whether the lunate sulcus (see Figure 15.7), a fissure on both sides of the occipital lobe that is involved with vision, was more ape- or human-like. The sulcus is smaller in humans than in monkeys and apes.
doi:10.15347/wjm/2014.010. ISSN 20018762.
The external skull reflected the cerebral expansion by becoming more rounded and exhibiting more of a forehead. In addition, the sagittal and pronounced nuchal muscle crests seen in Au. afarensis were not present. However, some males had convergent temporal lines that suggest that they may have had a slight sagittal crest. There is debate over whether the cranial base was flexed (see Figure 15.8), or whether that was a development seen only in more derived forms of Homo.
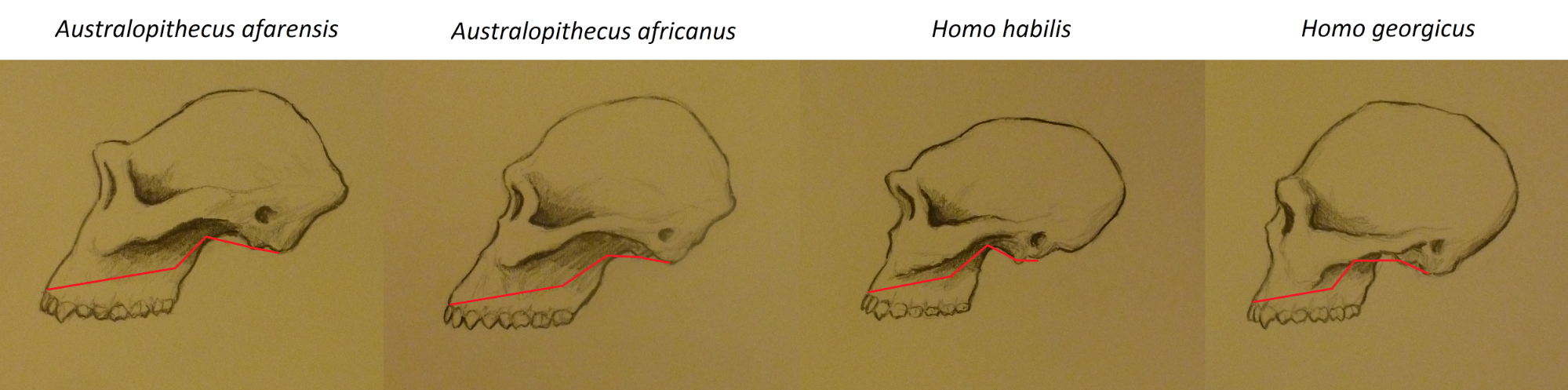
The species’ face was prognathic with a distinctive concavity in the midfacial region (see Figure 15.9). The dental arcade was more parabolic, and the teeth were smaller than those of Au. afarensis. The first premolar is considered to have been bicuspid, versus semi-sectorial in Au. afarensis. However, their faces were more heavily buttressed for chewing a tougher diet, and some researchers thus consider them to be “robust” australopiths.

They retained the primitive condition of long arms and their finger bones were somewhat curved. However, their hands were more human-like and they possessed our “power” thumb, giving them increased pinching and gripping capabilities. This was accomplished via better developed intrinsic thumb muscles, i.e. within the hand versus coming from the forearm to act on the thumb, and a specialized first metacarpal.
While they were considered to be habitual and very likely obligate bipeds, they still retained a divergent hallux. According to McHenry and Berger (1998) and Green et al. (2007) they may have been more arboreal than Au. afarensis.
Au. africanus were less sexually dimorphic than Au. afarensis, with males averaging 4′6″ (138 cm) tall and 90 lb (41 kg) and females at 3′9″ (115 cm) and 67 lb (29 kg).
Review of Primitive Characteristics
- Somewhat prognathic.
- Arboreal characteristics:
- Upward-oriented shoulder joints.
- Long arms relative to legs.
- Long, curved metacarpals and phalanges.
Review of Derived Characteristics
- Skull changes:
- More rounded skull vault.
- Increased encephalization.
- Frontal and parietal expansion.
- More of a forehead.
- Loss of sagittal crest and less pronounced nuchal crest.
- Facial buttressing.
- More of a parabolic dental arcade and reduced anterior dentition.
- Power thumb:
- Well-developed intrinsic thumb muscles.
- Specialized first metacarpal.
- Reduced sexual dimorphism.
ENVIRONMENT AND WAY OF LIFE
While there is some controversy as to how much time Au. africanus spent in open versus closed (forest) environments, their facial robusticity and molars reflect a greater reliance on tougher, more dry-adapted plant foods than earlier hominins. They retained climbing adaptations and thus could have used trees for safety, sleeping, and food. When on the ground, it is thought that they were scavenger-foragers, foraging for plant foods and small prey and scavenging remains of carnivore kills. Crude stone tools have been found at the sites of Sterkfontein and Makapansgat, and while there is no evidence of tool manufacture, it appears that they were using stones for hammering and cutting. They are thought to have relied on tubers for part of their diet and they likely used digging sticks to unearth them, in much the same way as modern African peoples, such as the San-speaking groups of the Kalahari Desert.
As mentioned, it is thought that hominins may have fallen through the surface into subterranean caves and/or bones were dragged in by rodents for gnawing. While it is possible that they used open caves for shelter from sun and rain, there is no evidence that they lived in caves.
In nature, extractive foragers have relatively large brains. Examples within the primate world include the aye-aye (a prosimian of Madagascar) that hunts insects via a combination of audition (hearing) and extraction (see www.arkive.org for multiple videos); New World capuchin monkeys that steal young animals from tree holes and rip bark from trees to get at insects; open-country baboons that dig for tubers; and the brainy orangutans and chimps that use a variety of tools to extract a range of foods, from ants to honey (see Figures 15.10 and 15.11 for videos of chimps and orangutan). Chimps have even been observed using sticks to spear bushbabies (small nocturnal African prosimians) in tree holes (e.g. see “How Smart Are Planet’s Apes?” on NationalGeographic.com). The same can be seen in some bird species, most notably tool-using crows and nut-burying nuthatches. Since our last common ancestor with chimps and bonobos was already relatively smart, I believe that while bipedalism was in response to diminishing habitat and resources, encephalization was in response to having to locate and learn to process new foods. Beginning with Au. africanus and the subsequent paranthropines, we see an upward trend for encephalization and expansion of those portions of the brain involved with association, i.e. complex thought processing, as well as increasing manual strength and dexterity for manipulating tools and objects. Thus we stepped out and then had to find and fix dinner!
We tend to think of “evolution by means of natural selection” as being a positive aspect of life, i.e. “survival of the fittest.” However, when there is a “rapid” response to a changing environment, many individuals die out while those few survive.
Videos
Ant dipping 1 by TheFriendsAndAi, https://youtu.be/ScqG54B4KtE
Orangutan using tool by Alain Compost, https://youtu.be/pXOTAgq-eiY

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}